Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
И овогенез
|
|
Половое размножение встречается как у одноклеточных, так и у многоклеточных растений и животных.
Как отмечено в главах II и III, половое размножение у бактерий осуществляется путем конъюгации, служащей аналогом полового процесса и являющейся системой рекомбинации этих организмов, тогда как у простейших половое размножение происходит тоже путем конъюгации либо путем сингамии и аутогамии.
У многоклеточных организмов (растений и животных) половое размножение связано с образованием зародошевых или половых клеток (гамет), оплодотворением и образованием зигот.
Половое размножение является значительным эволюционным приобретением организмов. С другой стороны, оно способствует пересортировке генов, появлению разнообразия организмов и повышению их конкурентоспособности в непрерывно меняющихся условиях окружающей среды.
У одноклеточных организмов половое размножение существует в нескольких формах. У бактерий половое размножение можно анало-гизировать с имеющими место у них конъюгацией, заключающейся в передаче плазмидной или хромосомной ДНК от клеток-доноров (содержащих плазмиды) к клеткам-реципиентам (не содержащих плазмиды), а также с трансдукцией бактерий, заключающейся в передаче генетического материала от одних бактериальных клеток к другим фагам. Конъюгация встречается также у инфузорий, у которых во время этого процесса происходит переход ядер от одних особей к другим, после чего следует деление последних.
Одной из распространенных форм полового размножения у одноклеточных животных, например, у паразитических споровиков, является копуляция. Половое размножение у них заключается в слиянии двух особей, которые являются гаметами, в одну, являющуюся споровой формой, из которой затем развивается новый организм. Независимо от способа слияния генетического материала у одноклеточных организмов существенной особенностью этого слияния является то, что оно сопровождается генетической рекомбинацией.
У многоклеточных растений и животных половое размножение происходит через образование женских и мужских половых клеток (яйцеклеток и сперматозоидов), последующее оплодотворение яйцеклетки сперматозоидом и образование зиготы. У растений половые клетки продуцируются в специализированных репродуктивных структурах, у животных они продуцируются в половых железах, называемых гонадами (от греч. gone — семя).
Между соматическими и половыми клетками животных существует важное различие. Оно заключается в том, что соматические клетки способны к делению, т. е. репродуцируют себя и кроме того из них образуются половые клетки. Напротив, половые клетки не делятся, но они «начинают» репродукцию целого организма.
Диплоидные соматические клетки, из которых образуются мужские половые клетки, называют сперматогониями, а диплоидные соматические клетки, из которых образуются женские половые клетки — овогониями. Процесс образования (роста и дифференциации) мужских и женских половых клеток носит название гаметогенеза.
Гаметогенез основан на мейозе (от греч. meiosis — понижать), который представляет собой процесс редукционного деления ядер клеток, сопровождающегося понижением числа хромосом на ядро. Мейоз происходит в специализированных клетках репродуктивных органов живых существ, размножающихся половым путем (рис. 78). Например, у папоротникообразных мейоз встречается в специализированных клетках спорангий, располагающихся на нижней поверхности листьев этих растении и развивающихся в споры, а затем в га-метофиты. Последние существуют раздельно, продуцируя в конечном итоге мужские и женские гаметы. У цветковых растений мейоз осуществляется в специализированных клетках семяпочек, которые развиваются в споры.
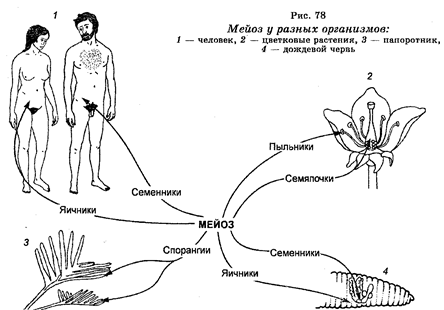
Последние продуцируют гаметофит с одной яйцеклеткой.
 Кроме того у этих растений мейоз происходит также в специализированных клетках пыльников, которые также развиваются в споры, продуцирующие в конечном итоге пыльцу с двумя мужскими гаметами. У земляных червей, которые являются гермафродитами и содержат мужские половые органы в одном сегменте тела, а женские — в другом и которые характеризуются способностью к перекрестному оплодотворению между разными особями, имеется способность одновременно к сперматогенезу и овогенезу.
Кроме того у этих растений мейоз происходит также в специализированных клетках пыльников, которые также развиваются в споры, продуцирующие в конечном итоге пыльцу с двумя мужскими гаметами. У земляных червей, которые являются гермафродитами и содержат мужские половые органы в одном сегменте тела, а женские — в другом и которые характеризуются способностью к перекрестному оплодотворению между разными особями, имеется способность одновременно к сперматогенезу и овогенезу.
У млекопитающих мейоз осуществляется в специализированных клетках семенников и яичников, в которых продуцируются мужские и женские гаметы соответственно. Выявлены белки — индукторы мейоза.
В процессе мейоза дипдоидное число хромосом (2п), которое характерно для соматических клеток (клеточных ядер) и незрелых зародышевых клеток, изменяется до гаплоидного числа (In), характерного для зрелых зародышевых клеток. Таким образом, в результате гаметогенеза половые клетки получают лишь половину хромосом соматических клеток (рис. 79).
Поведение хромосом в период гаметогенеза у животных является одинаковым как у мужских, так и у женских особей. Однако пол различается во времени происхождения разных стадий мейоза, что особенно заметно у человека. У мужских особей в постпубертатный период полный процесс мейоза завершается примерно в течение двух месяцев, в то время как у женских особей первое мейотическое деление начинается еще в фетальном яичнике и не завершается, пока не начнется овуляция, которая наступает у них примерно в пятнадцать лет.
У высших животных в случае мужских особей мейоз сопровождается образованием четырех функционально активных гамет (рис. 80). Напротив, у женских особей каждый овоцит II порядка дает лишь одну яйцеклетку. Другие ядерные продукты женского мейоза представляют собой три редукционных тельца, которые не участвуют в размножении и дегенерируют.
Мейоз состоит из двух делений клеточного ядра, которые называют мейотическими. Первое мейотическое деление ядра разделяет членов каждой пары гомологичных хромосом после того, как они спарились одна с другой (синапсис) и обменялись генетическим материалом (кроссинговер). В результате этого разделения образуется два гаплоидных ядра. Второе мейотическое деление разделяет две продольные половины хромосом (хроматиды) в каждом из этих ядер, продуцируя четыре гаплоидных ядра.
В процессе гаметогенеза происходит также дифференциация яйцеклеток (овогенез) и сперматозоидов (сперматогенез), являющаяся пререквизитом их функций. Будучи высокоспециализированными структурами, яйцеклетки животных намного крупнее сперматозоидов, обычно неподвижны и содержат питательный материал, который обеспечивает развитие эмбриона в начальном периоде после оплодотворения. Сперматозоиды большинства животных обладают жгутиком, обеспечивающим независимость их движения.

Мейоз имеет выдающееся биологическое значение. Благодаря мейозу в клетках организмов поддерживается постоянное число хромосом независимо от количества поколений. Следовательно, мейоз поддерживает постоянство видов. Наконец, в мейозе в результате кроссинговера происходит рекомбинация генов, которая является одним из факторов эволюции, хотя его значение является и меньшим по сравнению с мутагенезом.
Сперматогенез — это процесс образования зрелых мужских половых клеток. Сперматозоиды развиваются в мужских половых железах (семенниках, или тестисах) из специализированных соматических клеток (рис. 81). Такими специализированными клетками служат так называемые примордиальные зародышевые клетки, которые мигрируют к тестисам в раннем периоде эмбриогенеза мужского индивидуума. Следовательно, примордиальные клетки являются прародителями (предшественниками) зрелых половых клеток. У млекопитающих после достижения половой зрелости сперматозоиды образуются практически всю жизнь.
Семенники человека состоят из многочисленных канальцев, стенки которых сформированы слоями клеток, которые находятся на разных стадиях развития сперматозоидов. Наружный слой канальцев образован крупными клетками, называемыми сдерма-тогониями. Эти клетки содержат диплоидный набор хромосом и являются в тестисах потомками примордиальных зародышевых клеток.

В период половой зрелости индивида часть сперматогоний перемещается во внутренний слой канальцев, где в результате мейоза они развиваются в клетки, получившие название сперматоцитов первого порядка (сперматоцитов I), затем в спер-матоциты второго порядка (сперматоциты II) и, наконец, в сперматиды, являющиеся гаплоидными зародышевыми клетками, дифференцирующимися в конечном итоге в зрелые сперматозоиды. Таким образом, в обобщенном виде можно сказать, что сперматогенез инициируется в диплоид-ных соматических клетках (сперматогони-ях), после чего следует период созревания зародышевых клеток, в котором происходит два деления ядер путем мейоза, приводящего к образованию сперматид.
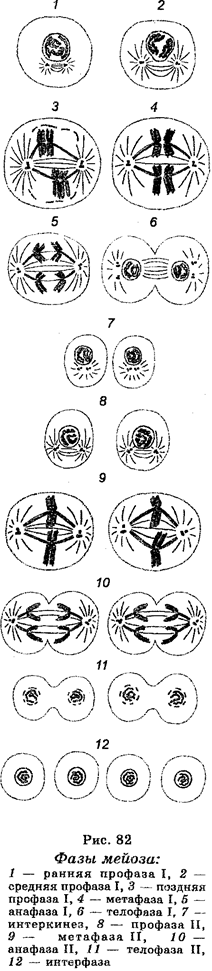 Мейоз в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I ® первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) ® интерфаза II (интерокинез) ® второе мейотическое деление (профаза II, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а скорее свойства хромосом на разных стадиях (рис. 82). Интерфаза I характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.
Мейоз в сперматогенезе протекает в несколько стадий (фаз). Между делениями имеются две интерфазы. Таким образом, мейотическое деление можно представить в виде серии следующих одно за другим событий, а именно: интерфаза I ® первое мейотическое деление (ранняя профаза I, поздняя профаза I, метафаза I, анафаза I, телофаза I) ® интерфаза II (интерокинез) ® второе мейотическое деление (профаза II, метафаза II, анафаза II, телофаза II). Процесс мейоза очень динамичен, поэтому микроскопические различия между разными стадиями отражают скорее не характер самих стадий, а скорее свойства хромосом на разных стадиях (рис. 82). Интерфаза I характеризуется тем, что в ней происходит репликация хромосом (удвоение ДНК), которая к началу ранней профазы I почти полностью завершается.
Первое мейотическое деление начинается в первичном сперматоците и характеризуется длинной профазой, которая состоит из переходящих одна в другую профазы I и профазы II. В профазе I различают пять главных стадий — лептонему, зигонему, пахинему, диплонему и диакинез.
На стадии лептонемы хромосомы в ядре представлены в виде тонких спирализованных нитей, содержащих многочисленные темноок-рашенные гранулы (хромомеры). Расщепления хромомер и нитей не отмечают, но считают, что хромосомы на уровне этой стадии являются двойными, т. е. диплоидными. Гомологи каждой хромосомной пары объединяются хромомерами вдоль их длины по принципу застежки.
На стадии зигонемы характерно установление синапсов между гомологичными хромосомами, в результате чего образуются спаренные хромосомы (биваленты). Хромосомы Х и Y ведут себя по сравнению с аутосомами несколько по-другому. Они конденсируются в темноокрашиваемые гетерохроматиновые тела, спариваемые в результате наличия гомологичных районов на их концах.
На стадии пахинемы, которая является наиболее долгой во времени стадией в мейотической профазе, происходит конденсация бивалентов и разделение каждой хроматиды надвое, в результате чего каждый бивалент представляет собой сложную спиральную структуру, состоящую из четырех сестринских хроматид (тетрад). В конце этой стадии начинается разделение спаренных хромосом-бивалентов. Теперь гомологичные хромосомы могут наблюдаться рядом. Поэтому в некоторых препаратах можно видеть четыре хромосомы, которые образуются в результате дупликации каждого гомолога, формирующего сестринские хроматиды. На этой стадии происходят обмены между гомологами и формирование хиазм.
На стадии диплонемы происходит укорачивание, утолщение и взаимное отталкивание сестринских хроматид, в результате чего хроматиды в биваленте почти разъединены. Разделение является неполным по той причине, что в каждой паре хромосом еще не расщеплена центромера. Что касается бивалентов, то они удерживаются на различных местах вдоль их длины с помощью хиазм, которые являются структурами, сформированными между гомологичными хроматидами в результате предыдущего кроссинговера между синаптически связанными гомологами. В хороших препаратах можно наблюдать от одной до нескольких хиазм в зависимости от длины бивалента. Каждая наблюдаемая на этой стадии хиазма представляет собой результат обмена, который встречался между несестринскими хроматидами в течение стадии пахинемы. Поскольку сжатие и отталкивание бивалентов усиливается, хиазмы двигаются к концам хромосом, т. е. происходит терминализа-ция хромосом. В конце диплонемы наступает деспирализация хромосом; гомологи продолжают отталкиваться друг от друга.
На стадии диакинеза, которая сходна с диплотеной, продолжается укорочение бивалентов и наступает ослабление (уменьшение) хиазм, вследствие чего формируются дискретные единицы в виде хроматид (четырех). Непосредственно после завершения этой стадии происходит растворение ядерной мембраны.
В метафазе I биваленты достигают наибольшей конденсации. Становясь овальными, они располагаются в экваториальной части ядра, где формируют экваториальные пластинки мейотической метафазы I. Форма каждого бивалента определяется числом и локализацией хиазм. У мужчин число хиазм на бивалент в метафазе I составляет обычно 1—5. Бивалент XY становится палочковидным в результате одиночной терминально расположенной хиазмы.
В анафазе I начинается движение противоположных центромер к противоположным полюсам клетки. В результате этого происходит разделение гомологичных хромосом. Каждая хромосома состоит теперь из двух хроматид, удерживаемых центромерой, которая не делится и остается интактной. Этим анафаза I мейоза отличается от анафазы митоза, при котором центромера подвергается разделению. Важно заметить, что благодаря кроссинговеру каждая хро-матида является генетически различной.
В стадии телофазы I хромосомы достигают полюсов, чем заканчивается первое мейотическое деление. После телофазы I наступает короткая интерфаза (интеркинез), в которой хромосомы деспи-рализуются и становятся диффузными, или телофаза I переходит прямо в профазу II второго мейотического деления. Ни в одном, ни в другом случае репликации ДНК не отмечается. После первого мейотического деления клетки называют спермат&цитами II порядка. Количество хромосом в каждой такой клетке снижается от 2п до п, но содержание ДНК еще не изменяется.
Второе мейотическое деление осуществляется в течение нескольких фаз (профаза II, метафаза II, анафаза II, телофаза II) и сходно с митотическим делением. В профазе II хромосомы вторичных спер-матоцитов остаются у полюсов. В метафазе П центромера каждой из двойных хромосом делится, обеспечивая каждую новую хромосому собственной центромерой. В анафазе II начинается формирование веретена, к полюсу которого двигаются новые хромосомы. В телофазе II второе мейотическое деление заканчивается, в результате чего каждый сперматоцит II порядка дает два спермати-да, из которых дифференцируются затем сперматозоиды. Как и во вторичном сперматоците, число хромосом в сперматиде является гаплоидным (п). Однако хромосомы сперматид являются одиночными, тогда как хромосомы вторичных сперматоцитов II являются двойными, будучи построенными из двух хроматид. Следовательно, ядро каждого сперматида имеет одиночный набор негомологичных хромосом. Вторичное мейотическое деление является делением митотического типа (экваториальным делением). Оно разделяет двойные сестринские хроматиды и отличается от редукционного деления, в котором гомологичные хромосомы разделены. Единственное существенное отличие от классического митоза заключается в том, что здесь имеется гаплоидный набор хромосом.
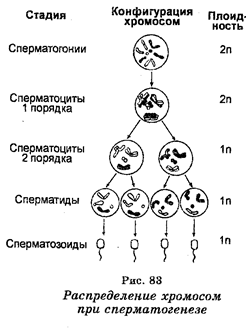 Итак, первое мейотическое деление сперматоцитов I порядка приводит к образованию двух вторичных сперматоцитов (II порядка). Обе хро-матиды структур, образующиеся в результате редукционного деления, являются сестринскими хроматида-ми. Последние возникают в результате репликации, предшествующей первому мейотическому делению. Второе мейотическое деление каждого вторичного сперматоцита приводит к образованию четырех сперма-тидов. Таким образом, в типичном мейозе клетки делятся дважды, тогда как хромосомы только один раз (рис. 83).
Итак, первое мейотическое деление сперматоцитов I порядка приводит к образованию двух вторичных сперматоцитов (II порядка). Обе хро-матиды структур, образующиеся в результате редукционного деления, являются сестринскими хроматида-ми. Последние возникают в результате репликации, предшествующей первому мейотическому делению. Второе мейотическое деление каждого вторичного сперматоцита приводит к образованию четырех сперма-тидов. Таким образом, в типичном мейозе клетки делятся дважды, тогда как хромосомы только один раз (рис. 83).
Конечный этап в сперматогенезе связан с дифференциацией, которая заканчивается тем, что каждый из сравнительно больших, сферических неподвижных сперматидов превращается в небольшой вытянутый подвижный сперматозоид.
 У большинства взрослых (сексуально зрелых) мужских особей животных сперматогенез происходит в семенниках постоянно или периодически (сезонно). Например, у насекомых для завершения цикла сперматогенеза требуется лишь несколько дней, тогда как у млекопитающих этот цикл затягивается на недели и даже месяцы. У взрослого человека сперматогенез проходит в течение всего года. Время развития примитивных сперматогоний В зрелые сперматозоиды составляет около 74 дней.
У большинства взрослых (сексуально зрелых) мужских особей животных сперматогенез происходит в семенниках постоянно или периодически (сезонно). Например, у насекомых для завершения цикла сперматогенеза требуется лишь несколько дней, тогда как у млекопитающих этот цикл затягивается на недели и даже месяцы. У взрослого человека сперматогенез проходит в течение всего года. Время развития примитивных сперматогоний В зрелые сперматозоиды составляет около 74 дней.
Мужские половые клетки, продуцируемые организмами разной видовой принадлежности, характеризуются подвижностью и чрезвычайным разнообразием в размерах и по структуре (рис. 84). Например, длина сперматозоидов D. melanogaster составляет 1,76 мм, что в 300 раз превышает длину сперматозоидов человека. Больше того, длина сперматозоидов D. bifurca составляет более 28 мм, что превышает длину самих насекомых этого вида в двадцать раз.
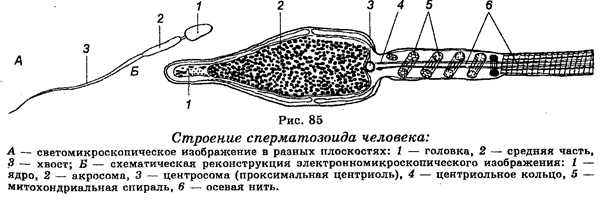
Каждый сперматозоид человека состоит из трех отделов — головки, средней части и хвоста (рис. 85). В головке сперматозоида располагается ядро. В нем содержится гаплоидный набор хромосом. Головка снабжена акросомой, которая содержит литические ферменты, необходимые сперматозоиду для вступления в яйцеклетку. В головке локализуется также две центриоли — прокси-мальная, которая побуждает деление оплодотворенной сперматозоидом яйцеклетки, и дистальная, которая дает начало аксимальному стержню хвоста. В средней части сперматозоида располагаются ба-зальное тельце хвоста и митохондрии. Хвост (отросток) сперматозоида образован внутренним аксимальным стержнем и внешним футляром, имеющим цитоплазматическое происхождение. Сперматозоиды человека характеризуются значительной подвижностью.
Овогенез — это процесс формирования яйцеклеток. Его функциями являются обеспечение гаплоидного набора хромосом в ядре яйцеклетки и обеспечение питательных потребностей зиготы. Овогенез в своем проявлении в основном сопоставим со сперматогенезом.
У млекопитающих и человека овогенез начинается еще во внутриутробном периоде (до рождения). Овогонии, представляющие собой мелкие клетки с довольно крупным ядром и локализованные в фолликулах яичников, начинают в фолликулах дифференцироваться в первичные овоциты. Последние формируются уже на третьем месяце внутриутробного развития, после чего они вступают в профазу первого мейотического деления. Ко времени рождения девочки все первичные овоциты уже находятся в профазе первого мейотического деления. Первичные овоциты остаются в профазе вплоть до наступления половой зрелости женского индивида. Когда при наступлении половой зрелости фолликулы яичника созревают, мейотическая профаза в первичных овоцитах возобновляется. Первое мейотическое деление для каждой развивающейся яйцеклетки завершается вскоре перед временем овуляции этой яйцеклетки. В результате первого мейотического деления и неравномерного распределения цитоплазмы одна образовавшаяся клетка становится вторичным овоцитом, другая — полярным (редукционным) тельцем.
Вторичное мейотическое деление у человека наступает, когда вторичный овоцит (развивающаяся яйцеклетка) проходит из яичника в фаллопиеву трубу. Однако это деление не завершается до тех пор, пока во вторичный овоцит не проникает ядерное содержимое сперматозоида, что происходит обычно в фаллопиевой трубе. Когда ядро сперматозоида проникает во вторичный овоцит, то последний делится, в результате чего образуется овотида (зрелая яйцеклетка) с пронуклеусом, содержащим одиночный набор из 23 материнских хромосом. У некоторых других видов образуются яйцеклетки, определяющие как мужской пол, так и женский. Важно подчеркнуть, что здесь происходит также расщепление и рекомбинация генов, основу которой создает расхождение хромосом. Другая клетка, образующаяся в результате второго мейотического деления у человека, является вторым полярным тельцем, не способным к дальнейшему развитию. В это время подвергается делению надвое и полярное (редукционное) тельце. Таким образом, развитие одного овоцита первого порядка сопровождается образованием одной овотиды и трех редукционных телец. В яичниках таким путем на протяжении жизни обычно созревает 300-400 ово-цитов, но в месяц созревает лишь один овоцит. В течение дифференциации яйцеклеток формируются мембраны, уменьшается в размере их ядро.
У некоторых видов животных овогенез протекает быстро и непрерывно и приводит к образованию большого количества яйцеклеток.
Вопреки сходству со сперматогенезом овогенез характеризуется некоторыми специфическими особенностями. Питательный материал (желток) первичного овоцита не распределяется поровну между четырьмя клетками, которые образуются в результате мей-отических делений. Основное количество желтка сохраняется в одной большой клетке, тогда как полярные тельца содержат очень мало этого вещества. Первые и вторые полярные тельца получают в результате делений те же самые хромосомные наборы, что и вторичные овоциты, но они не становятся половыми клетками. Поэтому яйцеклетки намного богаче питательным материалом по сравнению со сперматозоидами. Особенно сильно это различие проявляется в случае яйцекладущих животных.
Яйцеклетки млекопитающих имеют овальную или несколько вытянутую форму (рис. 86) и характеризуются типичными чертами клеточного строения. Они содержат все структуры, характер  ные для соматических клеток, однако внутриклеточная организация яйцеклетки очень специфична и определяется тем, что яйцеклетка является также и средой, которая обеспечивает развитие зиготы. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. У очень многих животных различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется еще на стадии овоцита. Представляя собой поверхностный слой овоцита, она
ные для соматических клеток, однако внутриклеточная организация яйцеклетки очень специфична и определяется тем, что яйцеклетка является также и средой, которая обеспечивает развитие зиготы. Одна из характерных особенностей яйцеклеток заключается в сложности строения их оболочек. У очень многих животных различают первичную, вторичную и третичную оболочки яйцеклеток. Первичная оболочка (внутренняя) формируется еще на стадии овоцита. Представляя собой поверхностный слой овоцита, она
имеет сложную структуру, т. к. пронизана выростами прилегающих к ней фолликулярных клеток. Вторичная (средняя) оболочка полностью сформирована фолликулярными клетками, а третичная (наружная) образована веществами, представляющими собой продукты секреции желез яйцеводов, через которые проходят яйцеклетки. У птиц, например, третичными оболочками яйцеклеток служат белковая, подскорлуповая и скорлуповая оболочки. Для яйцеклеток млекопитающих характерно наличие двух оболочек. Структура внутриклеточных компонентов яйцеклеток специфична в видовом отношении, а иногда имеет даже индивидуальные особенности.
Date: 2015-09-25; view: 708; Нарушение авторских прав