Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Сцепленное наследование
|
|
Г. Мендель проследил наследование семи пар признаков у гороха. Многие исследователи, повторяя опыты Менделя, подтвердили открытые им законы. Было признано, что эти законы носят всеобщий характер. Однако в 1906 г. английские генетики В.Бэтсон и Р.Пеннет, проводя скрещивание растений душистого горошка и анализируя наследование формы пыльцы и окраски цветков, обнаружили, что эти признаки не дают независимого распределения в потомстве. Потомки всегда повторяли признаки родительских форм. Стало ясно, что не для всех генов характерно независимое распределение в потомстве и свободное комбинирование.
Каждый организм имеет огромное количество признаков, а число хромосом невелико. Следовательно, каждая хромосома несет не один ген, а целую группу генов, отвечающих за развитие разных признаков.
| |||
| |||
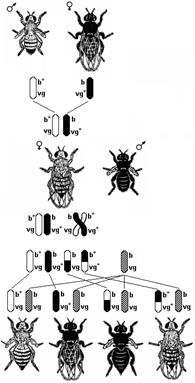 Рис. 327. Явление сцепленного
наследования признаков
у мушки дрозофилы.
Рис. 327. Явление сцепленного
наследования признаков
у мушки дрозофилы.
 Томас Гент Морган
(1886 — 1945)
Томас Гент Морган
(1886 — 1945)
Изучением наследования признаков, гены которых локализованы в одной хромосоме, занимался выдающийся американский генетик Т. Морган. Если Мендель проводил свои опыты на горохе, то для Моргана основным объектом стала плодовая мушка дрозофила. Мушка каждые две недели при температуре 25°С дает многочисленное потомство. Самец и самка внешне хорошо различимы — у самца брюшко меньше и темнее.
Кроме того, они имеют всего 8 хромосом в диплоидном наборе и отличия по многочисленным признакам, могут размножаться в пробирках на дешевой питательной среде.
Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибридов, имеющих серое тело и нормальные крыльяи (ген, определяющий серую окраску брюшка, доминирует над темной окраской, а ген, обусловливающий развитие нормальных крыльев — над геном недоразвитых) (рис. 327). При проведении анализирующего скрещивания самки F1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1:1:1:1. Однако в потомстве явно преобладали особи с признаками родительских форм (41,5% серые длиннокрылые и 41,5% черные с зачаточными крыльями) и лишь незначительная часть мушек имела перекомбинированные признаки (8,5% черные длиннокрылые и 8,5% серые с зачаточными крыльями).
Анализируя полученные результаты, Морган пришел к выводу, что гены, обусловливающие развитие серой окраски тела и длинных крыльев, локализованы в одной хромосоме, а гены, обусловливающие развитие черной окраски тела и зачаточных крыльев, — в другой. Явление совместного наследования признаков Морган назвал сцеплением. Материальной основой сцепления генов является хромосома. Гены, локализованные в одной хромосоме, наследуются совместно и образуют одну группу сцепления. Поскольку гомологичные хромосомы имеют одинаковый набор генов, количество групп сцепления равно гаплоидному набору хромосом (например, у человека 46 хромосом, или 23 пары гомологичных хромосом, соответственно количество групп сцепления в соматических клетках человека — 23). Явление совместного наследования генов, локализованных в одной хромосоме, называют сцепленным наследованием. Сцепленное наследование генов, локализованных в одной хромосоме, называют законом Моргана.
Вернемся к нашему примеру скрещивания мушек дрозофил. Если гены окраски тела и формы крыльев локализованы в одной хромосоме, то при данном скрещивании должны были получиться две группы особей, повторяющие признаки родительских форм, так как материнский организм должен образовывать гаметы только двух типов — АВ и ав, а отцовский — один тип — ав. Следовательно, в потомстве должны образовываться две группы особей, имеющих генотип ААВВ и аавв. Однако в потомстве появляются особи (пусть и в незначительном количестве) с перекомбинированными признаками, то есть имеющие генотип Аавв и ааВв. Каковы причины появления таких особей? Для объяснения этого факта необходимо вспомнить механизм образования половых клеток — мейоз. В профазе первого мейотического деления гомологичные хромосомы конъюгируют, и в этот момент между ними может произойти обмен участками. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Ав и аВ, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но поскольку кроссинговер происходит не во всех гаметах, числовое соотношение фенотипов не соответствует соотношению 1:1:1:1.
В зависимости от особенностей образования гамет, различают:
©
|

©
|

Соответственно этому различают:
© рекомбинантные (кроссоверные) особи — особи, возникшие с участием кроссоверных гамет;
© нерекомбинантные (некроссоверные) особи — особи, возникшие без участия кроссоверных гамет.
Гены в хромосомах имеют разную силу сцепления. Сцепление генов может быть:
© полным, если между генами, относящимися к одной группе сцепления, рекомбинация невозможна (у самцов дрозофилы полное сцепление генов, хотя у подавляющего большинства других видов кроссинговер протекает сходно как у самцов, так и у самок);
© неполным, если между генами, относящимися к одной группе сцепления, возможна рекомбинация.
Вероятность возникновения перекреста между генами зависит от их расположения в хромосоме: чем дальше друг от друга расположены гены, тем выше вероятность перекреста между ними. За единицу расстояния между генами, находящимися в одной хромосоме, принят 1 % кроссинговера. Его величина зависит от силы сцепления между генами и соответствует проценту рекомбинантных особей от общего числа потомков, полученных при скрещивании. Например, в рассмотренном выше анализирующем скрещивании получено 17% особей с перекомбинированными признаками. Следовательно, расстояние между генами серой окраски тела и длинных крыльев (а также черной окраски тела и зачаточных крыльев) равно 17%. В честь Т. Моргана единица расстояния между генами названа морганидой.
Результатом исследований Т.Моргана стало создание им хромосомной теории наследственности:
© гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален;
© каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены;
© гены расположены в хромосомах в определенной линейной последовательности;
© гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;
© сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинатных хромосом; частота кроссинговера:
¨ является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость);
¨ зависит от силы сцепления между генами: чем сильнее сцеплены гены, тем меньше величина кроссинговера (обратная зависимость);
© каждый вид имеет характерный только для него набор хромосом — кариотип.
40.4. Генетика пола
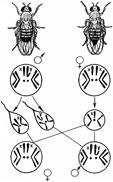 Рис. 328. Хромосомное определение пола у дрозофилы.
Рис. 328. Хромосомное определение пола у дрозофилы.
Как известно, большинство животных и двудомных растений являются раздельнополыми организмами, причем внутри вида количество особей мужского пола приблизительно равно количеству особей женского пола.
Пол можно рассматривать как один из признаков организма. Наследование признаков организма, как правило, определяется генами. Механизм же определения пола имеет иной характер — хромосомный (рис. 328).
Пол чаще всего определяется в момент оплодотворения. У человека женский пол является гомогаметным, то есть все яйцеклетки несут Х-хромосому. Мужской организм — гетерогаметен, то есть образует два типа гамет — 50% гамет несет Х-хромосому и 50% — Y-хромосому. Если
образуется зигота, несущая две Х-хромосомы, то из нее будет формироваться женский организм, если Х-хромосому и Y-хромосому — мужской.
|
 Рис. 329. Основные типы хромосомного определения пола.
Рис. 329. Основные типы хромосомного определения пола.
Из приведенной схемы видно, как происходит формирование в равных количествах двух групп особей, отличающихся набором половых хромосом.
Существует четыре основных типа хромосомного определения пола (рис. 329):
© мужской пол гетерогаметен; 50% гамет несут Х-, 50% —У-хромосому;
© мужской пол гетерогаметен; 50% гамет несут Х-, 50% —не имеют половой хромосомы;
© женский пол гетерогаметен; 50% гамет несут Х-, 50% —У-хромосому;
© женский пол гетерогаметен; 50% гамет несут Х-, 50% — не имеют половой хромосомы.
40.5. Наследование признаков,
сцепленных с полом
Генетические исследования установили, что половые хромосомы отвечают не только за определение пола организма — они, как и аутосомы, содержат гены, контролирующие развитие определенных признаков.
Наследование признаков, гены которых локализованы в Х- или Y-хромосомах, называют наследованием, сцепленным с полом.
Изучением наследования генов, локализованных в половых хромосомах, занимался Т.Морган.
У дрозофилы красный цвет глаз доминирует над белым. Проводя реципрокное скрещивание[41], Т.Морган получил весьма интересные результаты. При скрещивании красноглазых самок с белоглазыми самцами, в первом поколении все потомство оказывалось красноглазым. Если скрестить между собой гибридов F1, то во втором поколении все самки оказываются красноглазыми, а у самцов происходит расщепление — 50% белоглазых и 50% красноглазых. Если же скрестить между собой белоглазых самок и красноглазых самцов, то в первом поколении все самки оказываются красноглазыми, а самцы белоглазыми. В F2 половина самок и самцов — красноглазые, половина — белоглазые.
Объяснить полученные результаты наблюдаемого расщепления по окраске глаз Т.Морган смог, только предположив, что ген, отвечающий за окраску глаз, локализован в Х-хромосоме, а Y-хромосома таких генов не содержит.
Таким образом, благодаря проведенным скрещиваниям, был сделан очень важный вывод: ген цвета глаз сцеплен с полом, то есть находится в Х-хромосоме.
|
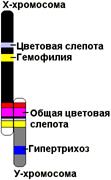 Рис. 330. Схема половых хромосом человека и сцепленных с ними генов.
Рис. 330. Схема половых хромосом человека и сцепленных с ними генов.
Х-хромосома человека содержит ряд генов, рецессивные аллели которых определяют развитие тяжелых аномалий (гемофилия, дальтонизм). Эти аномалии чаще встречаются у мужчин (так как они гетерогаметны), хотя носителем этих аномалий чаще бывает женщина.
У большинства организмов генетически активна только Х-хромосома, в то время как Y-хромосома практически инертна, так как не содержит генов, определяющих признаков организма. У человека лишь некоторые гены, не являющиеся жизненно важными, локализованы в Y-хромосоме (например, гипертрихоз — повышенная волосатость ушной раковины). Гены, локализованные в Y-хромосоме, наследуются особым образом — только от отца к сыну.
Полное сцепление с полом наблюдается лишь в том случае, если Y-хромосома генетически инертна. Если же в Y-хромосоме имеются гены, аллельные генам Х-хромосомы, характер наследования признаков иной. Например, если мать имеет рецессивные гены, а отец доминантные, то все потомки первого поколения будут гетерозиготны с доминантным проявлением признака. В следующем поколении получится обычное расщепление 3:1, причем с рецессивными признаками будут только девочки. Такой тип наследования называют частично сцепленным с полом. Так наследуются некоторые признаки человека (общая цветовая слепота, кожный рак).
40.6. Генотип целостная,
исторически сложившаяся система генов.
Изучая закономерности наследования, Г.Мендель исходил из предположения, что один ген отвечает за развитие только одного признака. Например, ген, отвечающий за развитие окраски семян гороха, не влияет на форму семян. Причем эти гены располагаются в разных хромосомах, и их наследование независимо друг от друга. Поэтому может сложиться впечатление, что генотип представляет собой простую совокупность генов организма. Однако сам Мендель в ряде опытов столкнулся с явлениями наследования, которые не могли быть объяснены с помощью открытых им закономерностей. Так, при изучении наследования окраски семенной кожуры, Мендель обнаружил, что ген, вызывающий образование бурой семенной кожуры, способствует также развитию пигмента и в других частях растения. Растения с бурой семенной кожурой имели цветки фиолетовой окраски, а растения с белой семенной кожурой — белые цветки. В других опытах, проводя скрещивание белой и пурпурной фасоли, он получил во втором поколении целый ряд оттенков — от пурпурного до белого. Мендель пришел к заключению, что наследование пурпурного цвета зависит не от одного, а от нескольких генов, каждый из которых дает промежуточную окраску. Можно говорить о том, что Мендель не только установил законы независимого наследования пар аллелей, но и заложил основы учения о взаимодействии генов.
После переоткрытия законов наследования признаков, многочисленные опыты подтвердили правильность установленных Менделем закономерностей. Вместе с тем, постепенно накапливались и факты, показывающие, что полученные Менделем числовые соотношения при расщеплении гибридного поколения не всегда соблюдались. Это указывало на то, что взаимоотношения между генами и признаками носят более сложный характер. Выяснилось, что:
© один и тот же ген может оказывать влияние на развитие нескольких признаков;
© один и тот же признак может развиваться под влиянием многих генов.
| Взаимодействие генов |
Как правило, взаимодействие генов имеет биохимическую природу, то есть оно основано на взаимодействии белков, синтезируемых под действием определенных генов. Взаимодействовать друг с другом могут как аллельные, так и неаллельные гены.
| Взаимодействие аллельных генов |
Различают несколько типов взаимодействия аллельных генов:
© Полное доминирование, при котором рецессивный признак не проявляется;
© Неполное доминирование, при котором у гибридов наблюдается промежуточный характер наследования.
© Кодоминирование, в этом случае у гибридов проявляются оба признака. Например, кодоминирование проявляется у людей с 4 группой крови. Первая группа крови у людей с аллелями iOiO, вторая — с аллелями IAIA или IAí0; третья — IВIВ или IВí0; четвертая группа имеет аллели IАIВ.
| Взаимодействие неаллельных генов |
Известно много примеров, когда гены влияют на характер проявления определенного неаллельного гена или на саму возможность проявления этого гена.
| Комплементарное взаимодействие |
Комплементарными называют гены, обусловливающие при совместном сочетании в генотипе в гомозиготном или гетерозиготном состоянии новое фенотипическое проявление признака.
|
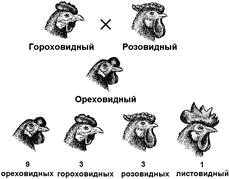 Рис. 331. Наследование формы гребня у кур.
Рис. 331. Наследование формы гребня у кур.
что куры с розовидным гребнем имеют генотип А_bb, с гороховидным — ааВ_, с ореховидным — А_В_ и с листовидным — ааbb, то есть развитие розовидного гребня происходит в том случае, если в генотипе имеется только один доминантный ген — А, гороховидного — наличие только гена В, сочетание генов А В обусловливает появление ореховидного гребня, а сочетание рецессивных аллелей этих генов — листовидного.
При комплементарном взаимодействии генов в дигибридном скрещивании получаются расщепления потомков отличные от менделевского: 9:7, 9:3:4, 13:3, 12:3:1, 15:1, 10:3:3, 9:6:1. Однако все они являются видоизменениями общей менделевской формулы 9:3:3:1.
| Эпистаз. |
Эпистатичным называют такое взаимодействие генов, при котором аллель одного гена подавляет действие аллелей других генов. Эпистатичное взаимодействие противоположно комплементарному. Некоторые породы кур имеют белое оперение, другие же — окрашенное.
|
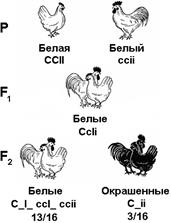 Рис. 332. Наследование окраски оперения у кур
Рис. 332. Наследование окраски оперения у кур
|
 Рис. 333. Наследование окраски зерновки у пшеницы.
Рис. 333. Наследование окраски зерновки у пшеницы.
При скрещивании, например, леггорнов (ССII)с плимутроками (ссii), все потомство F1 имеет белую окраску, которая определяется наличием в их генотипе гена-подавителя (СсIi). Если же гибридов F1 скрестить между собой, то во втором поколении происходит расщепление по окраске в отношении 13/16 белых: 3/16 окрашенных. Окрашенным оказывается та часть потомства, в генотипе которой имеется ген окраски и отсутствует его подавитель (С_ii).
| Полимерия |
Скрещивая белую и пурпурную фасоли, Мендель столкнулся с явлением полимерии. Полимерией называют однозначное влияние двух, трех и более неаллельных генов на разви-
тие одного и того же признака. Такие гены называют полимерными, или множественными, и обозначают одной буквой с соответствующим индексом, например, А1, А2, а1, а2.
Полимерные гены контролируют большинство оличественных признаков организмов: высоту растения, массу семян, масличность семян, содержание сахара в корнеплодах сахарной свеклы, удойность коров, яйценоскость, вес тела и т.д.
Явление полимерии было открыто в 1908 г. при изучении окраски зерновки у пшеницы Нельсоном-Эле (рис. 333). Он предположил, что наследование окраски у зерновки пшеницы обусловлено двумя или тремя парами полимерных генов. При скрещивании краснозерной и белозерной пшеницы в F1 наблюдалось промежуточное наследование признака: все гибриды первого поколения имели светло-красное зерно. В F2 происходило расщепление в отношении 63 краснозерных на 1 белозерное. Причем краснозерные зерновки имели разную интенсивность окраски — от темно-красной до светло-красной. Исходя из наблюдений, Нельсоном-Эле определил, что признак окраски зерновок обуславливает три пары полимерных генов.
У человека по типу полимерии наследуется, например, окраска кожи.
| Плейотропия |
Плейотропией называют множественное действие генов. Плейотропное действие генов имеет биохимическую природу: один белок-фермент, образующийся под контролем одного гена, определяет не только развитие данного признака,нои воздействует на вторичные реакции биосинтеза различных других признаков и свойств, вызывая их изменение.
Плейотропное действие генов впервые было обнаружено Г. Менделем, который обнаружил, что у растений с пурпурными цветками всегда имелись красные пятна в пазухах листьев, а семенная кожура была серого или бурого цвета. То есть развитие этих признаков определяется действием одного наследственного фактора (гена).
У человека встречается рецессивная наследственная болезнь—серповидно-клеточная анемия. Первичным дефектом этой болезни является замена одной из аминокислот в молекуле гемоглобина, что приводит к изменению формы эритроцитов. Одновременно с этим возникают глубокие нарушения в сердечно-сосудистой, нервной, пищеварительной, выделительной системах. Это приводит к тому, что гомозиготный по этому заболеванию погибает в детстве.
Плейотропия широко распространена. Изучение действия генов показало, что плейотропным эффектом, очевидно, обладают многие, если не все, гены.
Таким образом, выражение «ген определяет развитие признака» в значительной степени условно, так как действие гена зависит от других генов — от генотипической среды. На проявление действия генов влияют и условия окружающей внешней среды. Следовательно, генотип является системой взаимодействующих генов.
Date: 2015-09-05; view: 713; Нарушение авторских прав