Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Лекция жоспары
|
|
1. Потенциал туралы түсінік
2. Биопотенциал түрлері: тыныштық және әсер потенциалдары
3. Гольдман-Ходжкин-Катц теңдеуі.
4. Әрекет потенциалының пайда болу механизмі.
5. Ходжкин-Хаксли теңдеуі.
6. Әрекет потенциалының таралуы.
Лекция мақсаты: биопотенциалдың түрлерін, пайда болу механизімін, оның математикалық өрнегін және әрекет потенциалының нерв талшығымен таралуын талдау.
Физикада потенциал термині әр түрлі өрістердің (электростатикалық, магниттік, гравитатциялық т.б.) энергиясын сипаттау үшін қолданылатын шама. Биологиялық мембранада электр өрісі бар, олай болса ол өрістің энергиясын да осы тәрізді бипотенциал деген шамамен сипаттаймыз. Көбіне потенциалдың шамасын қолданбайды, олардың айырымын, яғни екі нүктедегі потенциалдар айрымын қолданады: Dj =j2 - j1, мұндағы j2 и j1 сәйкес нүктелердегі (аймақтардағы) потенциал шамасы. Биопотенциал деп ағзаның екі нүке арасындағы потенциалдар айырымын немесе мембранадағы биопотенциал деп, мембрананың ішкі және сыртқы орта арасындағы потенциалдар айырымын атайды: DjМ =jІШ - jСЫРТ.
Барлық уақытта қозбаған жасушада потенциал айырымы кездеседі, оны тыныштық потенциалы деп атайды, жасуша ішінде потенциал «теріс», сыртында «оң» таңбалы болады.
Биопотенциалдың пайда болу механизімін толық талдау үшін, біз алдымен олардың негізінде қандай физика-химиялық құбылыстардың жатқандығын, және потенциалдың шамасы неге байланысты болатындығын талдайық.
Пайда болу механизміне байланысты потенциалдар: диффузиялық, фазалық және мембраналық деген түрлерге бөлінеді. Диффузиялық потенциал сұйық ортаны бөліп тұрған шекарадағы иондардың қозғалғыштығының әр болуынан пайда болады (1 сурет).
HCl Н2О j- j+
Н+
Cl-
1 cурет
Мысалы, ортасында поры бар бөгет арқылы екіге бөлінген ыдыстың сол жағына тұз қышқылы ертіндісін (HCl), оң жағына су құяйық. Пор тек иондарды өткізсін делік. Диффузия құбылысы әсерінен және екі ортада концентрациялық градиенттің болуы себебінен иондар ыдыстың екінші жағына қарай қозғалады. Концентрациялық градиент деп белгілі бір бағыттағы бірлік ұзындыққа сәйкес келетін концентрация айрымына тең шаманы атайды. Мысалы, ертіндінің бірінші нүктедегі концентрациясы С1, одан l қашықтықта жатқан екінші нүктедегі концентрациясы С2 болса, онда gradC=С1- С2/ l тең болады.
Сонымен концентрациялық градиенттің нолден өзгеше болуы және оң зарядталған сутегі иондарының қозғалғыштығының жоғары болуы себепті, оң зарядталған сутегі иондары теріс зарядталған хлор иондарын артқа тастап, ыдыстың оң жағына бұрын жетеді, нәтижесінде ыдыстың оң жақ бөлігі оң зарядталады, ал сол жақ бөлігінде оң зарядтардың аз болуы (сутегі иондарының біраз бөлігі оң жаққа ауысып кетті) теріс зарядтардың басым болуы себепті ол орта теріс зарядталады. Осылайша ыдыстың екі жағындағы екі түрлі зарядтардың арасында электр өрісінің айырымы туындайды оны диффузиялық потенциал деп атаймыз және оның шамасы Гендерсон теңдеуімен анықталынады:

мұндағы u- катионның жылдамдағы, v- анионның жылдамдығы, R- газ тұрақтысы, T- абсолюттік температура, n- ионның валенттілігі, F- Фарадей саны, c1 – бірінші ортадағы ионның концентрациясы, c2 – екініш ортадағы ионның концентрациясы. Бұл өрнектен, дифффузиялық потенциалдың шамасы катион мен анионның қозғалғыштығының айрымашылығына, олардың концентрациясына тікелей байланысты екендігі көрінеді. Егер катион мен анионның жылдамдығы тең болса, немесе олардың концентрациясы бірде болса, онда диффузиялық потенциал нолге тең болады.
Биологиялық жүйелерде потенциалдың бүл түрі негізінен жасушаның механикалық зақымдалуы кезінде байқалады және иондардың зақымдалған аймақтан зақымдалмаған аймаққа қарай диффузиясы жүреді.
Қарастырылған мысалдағы бөгет аниондарды тіптен өткізбесін делік, сонда ыдыстың оң жағына тек сутегі иондары өтеді, яғни v =0 болғандықтан
Гендерсон теңдеуі Нернст теңдеуіне айналады: j= RT/nF ln C1/C2.
Фазалық потенциалдар араласпайтын екі фазалық күйдегі заттар шекарасында (судағы электролит ертіндісі мен май) пайда болады. Мысалы, анионға салыстырғанда катиондар майлы ортада жақсы еритін болса, онда ол майлы ортадаға жылдам ауысып, ортаны оң зарядтайды.
Мембраналық потециалдың пайда болуы туралы алғашқы теорияны 1902 жылы Бернштейн ұсынды. ХХ ғасырдың 50 жылдары Ходжикин, Хакси және Катц, Бернштейнннің теориясының дұрыстығын тәжірибе арқылы дәлелдеді. Олар мембрананың иондарды таңдап өткізуінен мембрананың іші мен сыртқы орталарындағы иондардың концентрациясының әр түрлі болуы биопотенциалдың пайда болуының басты себебі деген қорытынды жасады және оны кальмардың аксонында жүргізген тәжрибелері арқылы толық дәлеледі. Тәжірибе қозбаған мембрананың (кальмар аксоны) калий иондарын жақсы, ал натрии және басқа иондарды нашар өткізетіндігін көрсетті. Оның үстіне калии иондарының концентрациясы жасуша ішінде сыртқы ортамен салыстырғанда 20-40 есе көп, олай болса, диффузия әсерінен калии иондары сыртқа қарай тасымалданады, ал сыртқы ортадағы иондар ішке ене алмайды, олар үшін мембрана өтімділігі өте төмен. Сонымен сыртқы ортаға өткен калии иондары ортаны оң зарядтайды, ішкі ортадаға оң зарядтардың біраз бөлігін калий алып кеткендіктен, ішкі орта теріс зарядты болады (ішкі ортадағы теріс зарядтар сыртқа өте алмайды, оң зарядтар азайды). Бұл үрдіс динамикалық тепе теңдік орнағанға дейін жүреді. Сонымен тыныштық потенциалы калий иондарының циоплазмадан сыртқы ортаға қарай бағытталған диффузия әсерінен деп қарастырсақ, онда оның шамасын Нернст
теңдеуінен анықтауға болады:
j= RT/nF ln C1/C2
мұндағы К1 жасушаның ішіндегі, К2 сыртындағы калии ионының концентрациясы. Бұл теңдеуді тепе теңдік потенциалы деп те атайды. Кальмар аксонындағы тыныштық потенциалының шамасы jМ = - 60мВ (2 сурет).
| 1 2 3 4 5 6 t, мс |
| j, мВ -20 -40 -60 |
jМ = - 60мВ
2 сурет
Кейінгі жүргізілген тәжірибелер мембрананың аз мөлшерде натрия мен хлор иондарын да өткізетіндігін көрсетті, олай болса тыныштық потенциалының пайда болуында K+, Na+ және Cl- иондарынының да қатысы бар. Қозбаған мембрананың K+, Na+ және Cl- иондары өткізу коэффиценті PK: PNa: PCl = 1: 0,04: 45, мұнан калий ионының натриға салыстырғанда өтімділігі 25 есе жоғары екендігі көрінеді, яғни тыныштық потенциалының пайда болуында калии мен натрии иондарының концентрациялары басты роль атқаратындығы келіп шығады.
1949 жылы Гольдман, Ходжкин және Катц тыныштық потенциалының басты теңдеуін ұсынды:
jM = -RT/F ln 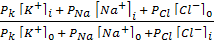 ,
,
мұндағы i и 0 иондардың ішкі және сыртқы ортадағы концентрациясы, РК, РNa РCl сәйкес иондар үшін мембрананың өткізгіштік коэффиценті. Бұл өрнек Гольдман- Ходжкин - Катц теңдеуі деп аталады.
Жасушаны сыртқы тітіркендіргіштер (механикалық, жылулық, электрлік) арқылы қозған күйге көшірсек, мембрананың натрии иондарын өткізуі күрт жоғарылайды, бірақ калии иондарын өткізу дәрежесі өзгеріссіз қалады. Мембрананың калии және натрии иондарын өткізу коэффициенті PK : PNa = 1: 20 дейін өзгеріп, натрии ионын өткізу 500 есе артады. Оның үстіне мембранада натрии иондарының каналы 10 есе көп, осы факторлар натрии иондарының мембрана ішіне қарай өтуін күшейтіп, ондағы потенциалдар айрымын төмендеуіне, соңынан оның ішкі қабатының оң зарядталуына алып келеді. Жасушадағы бұл құбылысты деполяризация деп атайды.
Мембрананың қозу нәтижесіндегі потенциалдар айрымының толық өзгерісі әрекет потенциалының мәнін береді: jӘ = jТ - Dj, где jТ – тыныштық потенциаланың мәні, Dj- потенциал өзгірісі. Әрекет потенциалы деп қозған күйдегі цитоплазма мен сыртқы орта арасындағы потенциалдар айрымын айтамыз. Мысалы, үлкен кальмар аксонының тыныштық потенциалы -60 Мв тең. Оны қоздырғанда потенцилы + 35 мВ дейін жоғарыласа, онда потенцилдың айырымы немесе әрекет потенциалы jӘ = - 60 мВ – 35 мВ = - 95 мВ (3- сурет).
| j,мВ -20 -40 -60 |
Dj
1 2 3 4 5 6 t, мс
jT jӘ
t
3- сурет. jТ – тыныштық потенциалы, jӘ – әрекет потенциалы, t- әсер потенциаланың ұзақтығы.
Ходжкин мен Хаксли мембранадағы кернеуді тұрақтандыру арқылы жүрізген тәжірибелерін қорытындылай келе, мембранадағы қозу құбылысының электрлік эквиваленті схемасын ұсынды (4 сурет).
| Сыртқы орта |
| Аксоплазма |
| RУТ IУТ jУТ |
| СМ |
RK RNa
IK
jK jNa
IK
4 сурет
Бұл модельге сәйкес қозған мембрана бір біріне параллель жалғанған С конденсатордан (мембраның сиымдылығына сәйкес), айнымалы екі кедергіден, олар калии gK = 1/RK және натрии gNa = 1/RNa каналдарының және үшінші кедергі хлор және басқа да иондардың gCl = 1/RCl өткізгіштігін сипатайды және осы тізбектегі тыныштық потенциалын сипаттайтын үш ЭҚК тұрады. Сонда қозған мембрана арқылы өтетін ток мына шамамен сипатталады: IM = CM dj/dt + IK + INa + I, мұндағы CM dj/dt - сиымдылықтоғы (мембрананың қайта зарядталуы кезіндегі ток шамсы), IK- калии иондарының тоғы, INa – натрии иондарының тогы, I – хлор және басқа да иондар тоғы. Мұндағы әр иондар тогы сол иондардың диффузиясы нәтижесінде пайда болатын тыныштық потенцила jМ мен тепе-теңдік Нернст потенциалнының jiP айрымына тең: Ii = gi(jM - jiP).
Осы айтылғандарды ескере келе Ходжикин мен Хаксли әсер потенциалы үшін мына түрдегі теңдеуді ұсынды:
IM = CM dj/dt + gK(jM - jKP) + gNa(jM - jNaP) + gУТ(jM - jУТP).
Бұл әрекет потенциалы үшін Ходжкин-Хаксли теңдеуі деп аталады және жүргізілген эксперимент нәтижелерімен жақсы үйлеседі.
Жасуша мембранасының қозу уақытының ұзақтығы өте аз, 0,5-1 мс аралығында жатады, онан соң мембрана қабатының натрии ионын өткізуі төмендейді, қайтадан тыныштық потенциалы орнайды. Бұл құбылыс реполяризация деп аталады. Әрекет потенциалының ұзақтығы жасуша түріне және температураға байланысты болады. Мысалы, температураны 100С кеміткенде әрекет потенциалының ұзақтығы үш есе артады.
Қозған мембранадағы әрекет потенциалы нерв талшығы (аксон) бойымен тарайды. Оның таралу механизмі төмендегіше: мембрананың қозған бөлігінің ішкі қабаты оң зарядталады (деполяризацияланады), ал қозбаған бөлігінің заряды (тыныштық потенциалы) теріс қалпында қалады, екі бөлік арасында потенциалдар айырымы пайда болып, ток жүреді, ол ток өткізгіш сыртқы қабат арқылы да жүріп тізбекті тұйықтайды (5- сурет).
| қозған. қозбаған. - - - - + + + + + + |
| Өткізгіш сыртқы орта |
| Өткізгіш сыртқы орта |
| Өткізгіш сыртқы орта |
+ + + + - - - - - - - - -
аксон
+ + + + - - - - - - - - -
- - - - - + + + + + + +
5- сурет.
Осылайша пайда болған «жергілікті (локальный) ток» деполяризацияланған бөлікке жанасып жатқан қозбаған нерв талшығының бөлігін тітіркендіріп, оны қоздырады, нәтижесінде ол бөлікте әрекет потенциалы пайда болады, бұл құбылыс келесі бөлікке ауысып, жалғаса береді. Ток жасушаның сыртқы қабаты арқылы қозбаған бөліктен қозған бөліке қарай жүреді, ал ішкі ортада ток керісінше бағытталады.
Кез келген электр тогы сияқты «жергілікті ток» мембрананы тіріркендіріп, мембрананың иондарды өткізуін жоғарылатады, бұл құбылыс өз кезегінде осы бөлікте әрекет потенциалын тудырады, ол келесі бөлікті қоздырады, процесс осылайша жалғасып, нерв талшығы бойымен импульс тарайды. Бұл кезде бұрын қозған мембрана бөлігінде қайта қалпына келу процессі, яғни реполяризация (тыныштық потенциланының орнауы) құбылысы жүреді. Импульстің аксон бойымен тарауы бір бағытта мембрананың қозған бөлігінен келесі бөлігіне қарай жүреді. Мембрананың осының алдында ғана қозуды өткізген бөлігі біраз уақыт бойы қозбаған - рефракторлы күйін сақтайды, сондықтан импульс кері бағытта тарамайды.
Нерв импулісінің (әрекет потенциалының) таралуы нерв талшығының қасиетіне байланысты. Нерв талшығы миелинді және миелинсіз деген екі түрге бөлінеді. Миелинді нерв талшығының цилиндр тәрізді орта бөлігі аксоплазмамен толтырылған, сырты цитоплазмалы мембранамен қапталған болып келеді. Нерв талшығының цилиндр тәрізді өсінің әр жерінің 0,2 мкм ден 2 мкм сайын қабаты жаңалаштанған болып келеді. Мұндай аймақтарды нерв талшығының түйіні немесе Ранвье үзілісі деп атайды және бұл аймақтар жасушаның сыртын алып жатқан ертіндімен жанасады. Ранвье үзіндісінің ұзындығы жобамен 1 мкм тең (6- сурет).
| Нерв талшағы |
| Сыртқы электр өткізгіш орта |
| Аксоплазма |
| Миелин |
| Ранвье үзілісі |
Сыртқы электр өткізгіш орта
6- сурет
Нерв жасаушасын миелинді қабаты липидтер мен ақуыздардан тұрады сондықтан ол изолятор болып табылады. Осы себепті қозу аксон мембранасының тек ашық аймақтарында (Ранвье үзіндісінде) пайда болады, бұл кезде аксонның басқа аймақтарында потенциалдар айырымы тұрақты, сыртқы орта оң, ішкі орта теріс зарядталған болып келеді. Миелинді нерв қабатынның деполяризацияланған А бөлігіндегі қозу келесі Б бөлігіне өтеді, бұл құбылыс ар қарай тек В нүктесінде жалғасады, ал бұрын қозған күйде болған Б нүктесі қозуды сезбейтін рефакторлы күйде біраз уақыт болады. Осы себепті импульстер нерв арқылы тек бір бағытта ғана тарайды (7- сурет).
| Сыртқы электр өткізгіш орта |
| В + |
| Б + |
| А - |
| + - - |
| Сыртқы электр өткізгіш орта Нерв импульсінің таралу бағыты |
7- сурет.
Миелинді нерв талшығы арқылы импульстердің осылайша таралуын секірмелі немесе сальтоторлы деп атайды. Импульстің сальтаторлы таралуы нерв импульстерінің таралу жылдамдығын артыруға мүмкіндік береді және мембрананың өте аз бөлігінің қозуы аз иондарды тасмалдауға, яғни натрии және калии иондық насостарының жұмысына аз энергия жұмсауға мүмкіндік туғызады.
Нерв талшығы арқылы әсер потенциалының х қашықтықа және t уақытқа байланысты таралуы телеграф теңдеуі арқылы сипатталады екен және оның шешуі мына өрнекке тең:
j = j0е-х/l,
мұндағы j - х нүктедегі потенциал мәні, х- қозған жерден потенциалы анықталынатын нүктеге дейінгі қашықтық, l - нерв талшығының тұрақтысы, ол деполяризия потенциалының е есе кемуіне тең ара қашықтық. Нерв талшығыынң тұрақтысы мына шамаға тең:
l = ÖrMDa/2ri,
мұндағы rM- нерв талшығы қабатының меншікті электр кедергісі, D - қабат қалыңдығы, a – нерв талшығының радиусы, ri – цитоплазманың меншікті электр кедергісі. Бұл өрнектерден миелинсіз нерв талшығы үшін нерв талшығы тұрақтысының үлкен шама болған сайын импульстің таралу жылдамдығы артатындығы байқалады, ал ол өз кезегінде аксон радиусы мен мембрананың меншікті кедергісінің үлкен, цитоплазманың меншікті кедергісінің аз болғандығын сайын үлкен шама болатындығын көреміз.
Миелинді нерв талшығында қозудың таралуы Ранвье үзіндісінің ұзындығына байланысты болады екен. Жүргізілген өлшеулер импульстің таралуы бір үзілістен екінші үзіліске дейін 0,07 мс екендігін көрсетті. Нерв радиусы 10 мкм болатын талшықтағы импульстардың таралу жылдамдығы 120 м/с тең.
Аутоиммундық аурулардың бірі склероз нерв талшығының миеленді қабаттың бұзылуынан, нерв талшығының ашылып қалуынан пайда болады. Соның әсерінен нерв импульстарының ашылып қалған нерв аймағындағы таралу жылдамдығы нашарлайды, нәтижесінде көру мен қозғалу қабілеті төмендейді, бұлшық еттердің әлсізденуі байқалады т.б. Сондай ақ- нерв талшығының демиелендігі басқа да аурулар себебінен де пайда болуы мүмкін, мысалы невралгия, радикулит, түрлі полиневропатология.
Қорытынды сұрақтар:
1. Потенциал қанадай физикалық шаманы сипаттайды?
2. Потенциалдың пайда болуын түсіндір.
3. Тыныштық потенциалының пайда болуы механизмі
4. Әрекет потенциалының пайда болуы механизмі
5. Миелинді және миленсіз нерв талшығында қозудың таралуы еркешелікктері
6. Ранвье үзіндісі.
7. Әрекет потенциалының таралуын түсіндір.
Date: 2015-09-19; view: 1314; Нарушение авторских прав