Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Эволюция
|
|
Археозой (4,55-2,5 млрд лет назад)
Согласно концепции абиогенеза (о химической эволюции жизни из неорганического материала), процесс начался в глубоководных гипертермальных источниках 4,2–4,0 млрд лет назад, а 4,0–3,7 млрд лет назад достиг поверхностных вод [525]. Жидкая вода возникла на Земле 4,3 млрд лет назад [575]. Следы ископаемых организмов появляются в палеонтологической летописи в позднем гадее св. 3,85 млрд лет назад [422], а также [156; 225; 282; 465; 574; 580; 600; 677; 678; 788]. Отметим, что по молекулярно-генетическим данным возраст первых организмов оценивается приблизительно в 1,8 млрд лет [578], что вдвое уступает палеонтологическому возрасту; по-видимому, 1,8 млрд лет назад – это возраст эволюционного расхождения тех организмов, чьи прямые потомки дожили до наших дней, поскольку молекулярно-генетический возраст, например, нескольких родственных видов живых существ определяется до момента разветвления эволюционных путей их предков.
Под влиянием мобилизующих колебаний продуктивности среды древние микроорганизмы развили пищевую автономность (автотрофность), чтобы не зависеть от внешних ресурсов в неурожайные (ледниковые) периоды. Около 3,8–3,5 (или 3,5; 3,465; 3,416) млрд лет назад цианобактерии освоили фотосинтез, т. е. способность синтезировать углеводы, аминокислоты, белки, пигменты и другие соединения под действием солнечного света [232; 788; 677; 678; 753]; а также [399; 465; 600]. Побочным продуктом фотосинтеза является свободный кислород. Однако поначалу он практически весь уходил на окисление железа, так что 2,8 млрд лет назад свободного кислорода в атмосфере почти не было [465, с. 2112]. Лишь 2,32 млрд лет назад наметился его атмосферный рост [181].
Чтобы добиться еще большей независимости от пищевых ресурсов среды, микроорганизмы путем симбиотического включения в свои тельца других одноклеточных существ обзавелись св. 2,8 млрд лет назад митохондриями [470], служащими внутренними источниками энергии для клеток, что подобно аккумуляторам повышало их мобильность. Митохондрии поначалу были самостоятельными микроорганизмами, а впоследствии поселились в тельцах других микроорганизмов и стали служить им «энергетическими подстанциями».
Примерно тогда же, св. 2,7 млрд лет назад, прокариоты (организмы без клеточного ядра) путем аналогичного симбиоза поселили в своих тельцах другие микроорганизмы, которые стали играть для них роль клеточных ядер. В результате прокариоты стали эукариотами, организмами с клеточными ядрами [213; 294; 322; 465; 718]. Этим способом содержащийся в клеточном ядре геном приобрел как бы скафандр в виде материнской клетки, что повысило его независимость от условий среды и ее перепадов, а следовательно подняло его мобильность. Поэтому хронологическая близость возникновения митохондрий и клеточных ядер может отвечать климатическому пессимуму, возможно связанному с оледенением, ознаменовавшим расхождение материков 2,85 млрд лет назад. Отметим, что в силу той же логики появление белковой оболочки (вириона) у РНК-вирусов тоже могло быть откликом на климатический или другой пессимум в их истории, однако палеонтология вирусов не развита.
Протерозой (2,5-0,544 млрд лет назад)
После распада позднеархейского суперконтинента Мегагеи, образовавшегося и расколовшегося 2,7–2,5 млрд лет назад, имели место три Гуронских ледниковых периода (Рамсей-Лейк, Брюс и Гоуганда) в интервале 2,5–2,2 млрд лет назад. Они, вероятно, связаны с эпизодом сильного расхождения материков (см. разд. 2.2). Падение продуктивности среды при III палеопротерозойском низкоширотном оледенении (III Гуронском) 2,222 ± 0,013 млрд лет назад [181; 313; 465] подстегнуло тягу жизни к самостоятельности вообще и автотрофности в частности. Поэтому жизнь ок. 2,2 млрд лет назад овладела способностью фиксировать азот [593], что ослабило ее зависимость от стационарных природных источников этого элемента, необходимого для поддержания существования живых организмов. Здесь мы имеем дело с одним из ранних фактов повышения мобильности организмов под влиянием оледенения, точнее под воздействием вызванной им убыли продуктивности среды.
Другим способом повысить мобильность организмов выступила многоклеточность. Современные бактерии способны образовывать колонии с высокоупорядоченным строением, в которых бактерии дифференцируются на клетки различных типов. Колонии некоторых миксобактерий (например Chondromyces crocatus) формируются в виде многоклеточного плодового тела со скоплениями одноклеточных спор на концах ответвлений. Это едва ли не прообраз многоклеточных растений, переживший свое время. Другие миксобактерии (например Myxococcus xanthus) образуют сферичные колонии из миллионов клеток. Такие колонии способны двигаться, захватывать и переваривать жертвы. Подобные хищные сферы в чем-то напоминают многоклеточных животных [115]. Многоклеточность последнего типа нацелена на подвижный сбор разреженной пищи, т. е. на ледниковые условия.
В соответствии с такими представлениями о назначении многоклеточности первые макроскопические многоклеточные водоросли появились в палеонтологической летописи 2,110 ± 0,052 млрд лет назад ([399; 698]) в Даспортскую ледниковую эпоху (2,2–1,95 млрд лет назад), вызванную образованием следующего сверхматерика (2,1–2,0 млрд лет назад). Наступившие вслед за Даспортским оледенением условия климатического оптимума расширили экологические ниши. Это произошло потому, что мягкие условия способствовали подъему биопродуктивности среды, а ее богатство сопровождалось ростом разнообразия природных условий и, соответственно, раздвижением экониш, что в свою очередь благоприятствовало умножению и экспансии жизни.
Судя по всему, простейшие организмы 1,2 млрд лет назад освоили новые для себя наземные водоемы [423]. Одновременно произошла крупная радиация (всплеск разнообразия) многоклеточных организмов, выразившаяся в разделении предков растений, грибов и животных [294; 322]; cp. [312; 780]. По молекулярно-генетическим данным, процесс продолжился отделением от животных губок, затем гребневиков, медуз и морских анемон, далее – плоских червей [312], а потом – дивергенцией (разделением) хордовых животных (к которым принадлежат и люди) и моллюсков (1,225 млрд лет назад), разделением хордовых и кольчатых червей (1,204 млрд лет назад), расхождением хордовых и членистоногих (1,173 млрд лет назад) и дивергенцией хордовых и иглокожих (1,001 млрд лет назад) [824]; cp. [294; 322; 470]. Дальнейший путь развития животных отмечен так называемой преэдиакарской биотой 980–900 млн лет назад [259, с. 220, рис. 1], которая связана с теплым эпизодом, благоприятным для умножения экониш и расцвета фауны.
Вслед за распадом еще одного сверхматерика возраста 1,1–1,0 млрд лет назад наступали ледниковые периоды Гнейсе, 900–850 млн лет назад, и Стёрт, 760–740 млн лет назад, отвечавшие состоянию разреженности материков. Очередной суперконтинент сформировался 675–600 млн лет назад, что повлекло за собой низкоширотное оледенение Марино-Варангер ок. 680–606 ± 3,7/ – 2,9, 602 ± млн лет назад [313; 381; 465; 807]. Низкоширотность (тропичность) оледенений Гурон III и Варангер, определенная палеомагнитным методом (когда по намагниченности ископаемых пород рассчитывают положение магнитных полюсов в древности), объясняется дрейфом магнитного полюса Земли относительно географического полюса. В протерозое этот дрейф превосходил современный и заметно искажал регистрацию палеоширот по намагниченности ископаемых пород.
Выгодность мобильности в ледниковых условиях оживила подвижных многоклеточных животных, представленных норами червей 620 млн лет назад [464; 682]. Последовавшее потепление эдиакарского периода (ок. 600–544 млн лет назад [312; 381]) создало множество экониш и вызвало дивергенцию бесчелюстных рыбообразных животных (круглоротых вроде миног и миксин) и челюстноротых (предков рыб) 599 млн лет назад [824; 479]. В результате развернулась богатая эдиакарская фауна возрастом в 565 ± 5–549/543 млн лет назад [203; 258; 381], отмеченная в финале своего расцвета молекулярно-генетической дивергенцией позвоночных животных (наших предков) 546 ± 18 млн лет назад [479; 838].
Палеозой (544-251 млн лет назад)
Развившееся хищничество одело мягкотелых эдиакарцев в твердые наружные покровы [258], поэтому кембрийский период (544–510/505 млн лет назад) ознаменовался широким распространением животных с защитным наружным скелетом, а также хордовых животных, чей внутренний скелет позволял создавать в воде локомоторную волну (толкающую животное вперед [124]) и повышал спасительную подвижность. Хищники производят эффект снижения биопродуктивности среды, поскольку мешают консументам (в данном случае – нехищным потребителям) беззаботно кормиться. Поэтому усвоение последними упомянутой спасительной подвижности типологически близко реакции на падение биопродуктивности среды вследствие похолодания. Следом за жертвами в локомоции усовершенствовались и хищники, примером которых может служить подвижный нижнекембрийский Anomalocaris (45 см), напоминавший плоскую сигару с боковыми плавниками и парой хватательных придатков возле рта. Организмы, отставшие от описанной тенденции, пришли к массовому вымиранию в раннем кембрии [184; 674, с. 431–437].
Целым «пакетом» в нижнем кембрии отмечены такие животные, как членистоногие [85, с. 41; 211; 239]; ракообразные [712]; иглокожие [700]; полухордовые (век атдабан, ок. 525 млн лет назад) [237; 701; 703; 727]; оболочники [699]; хордовые (век ботом) [237]; черепноподобные (530 млн лет назад) [238; 308]; миногообразные и миксинообразные (ок. 530 млн лет назад) [702; 838]; в верхнем кембрии – конодонты (родня позвоночных; 515 млн лет назад) [669; 210; 325; 437] и другие позвоночные (514 млн лет назад) [479]. Так выглядел древнейший эволюционный взрыв в истории Земли, обусловленный теплом и высокой биопродуктивностью эдиакарского и кембрийского периодов.
В следующем, ордовикском, периоде (510/505–438 млн лет назад) материки максимально отдалились друг от друга, что вызвало в позднем ордовике (458–438 млн лет назад) оледенение, условия которого обострили спрос на мобильность животных. Из числа головоногих моллюсков, появившихся в нижнем кембрии (например, наутилоидеи, потомки которых из рода Nautilus, живые ископаемые, и поныне здравствуют на юго-западе Тихого океана), особый расцвет в ордовике испытали представители подкласса эндоцератит [38, с. 356–359; 83, с. 173–204]. Их тело помещалось в длинно– и короткоконических раковинах прямых очертаний, игравших роль гидродинамических обтекателей. В раковине имелось образование – сифон, выполнявший гидростатические функции (он имелся и у других раковинных головоногих моллюсков и работал, как плавательный пузырь рыб, т. е. помогал своему обладателю менять общий удельный вес тела). Гидродинамическая раковина вместе с гидростатическим сифоном (он наполнял соответствующие камеры раковины азотом, аргоном и жидкостью, как у современного наутилуса, и тем самым регулировал удельный вес тела и его частей) делали эндоцератит идеальными пловцами, т. е. высокомобильными животными, типологически напоминающими по устройству подводные лодки на реактивной тяге.
В связи с похолоданием, поощряющим мобильность, эндоцератиты расцвели, стали владыками морей и породили самых крупных среди известных раковинных беспозвоночных, до 9,5 м длиной [38, с. 356; 83, с. 191]. Казалось бы, такие успешные монстры должны были бы процветать долго, однако с концом позднеордовикского оледенения они неизбежно пришли в упадок. Представители ведущего отряда, эндоцератиды, просуществовали весь ордовик и вымерли – возможно, один род данного отряда (Humeoceras) дожил до середины силура (421 млн лет назад) [38, с. 358]. Парадокс этот вполне объясним, если учесть, что высокомобильные формы вроде эндоцератит были узко приспособлены к скромной экосреде окололедникового времени и лишь в ней были сильны, а с подъемом биопродуктивности среды в силуре перестали ей соответствовать и угасли.
Прохладный ордовик был периодом радиации жизни [297], в особенности холодолюбивой – точнее, годной к низкой биопродуктивности среды. Холодолюбивые существа активно замещали ушедших в небытие теплолюбивых хозяев кембрия. Давление в пользу мобильных организмов привело на сушу наземную флору. Среди пионеров были наземные грибы (конец нижнего ордовика, ок. 460 млн лет назад), которые, по молекулярно-генетическим данным, возникли еще в конце Варангского оледенения ок. 600 млн лет назад [636], т. е. уже были испытаны суровыми условиями. Бок о бок с ними по суше распространялись наземные растения, отмеченные как в нижнем ордовике (475 млн лет назад), так и в верхнем (458–438 млн лет назад) [346, с. 481; 794]. В морях, судя по молекулярно-генетическим данным, появились подвижные лучеперые рыбы (450±35,5 млн лет назад) [86, с. 323–484; 479], а также акулы [468]. Лучеперые рыбы и акулы составляют свыше 95% современной ихтиофауны, захватившей моря благодаря кайнозойскому похолоданию. Их успех в современных условиях объясняется происхождением из позднеордовикского оледенения (см. далее).
Акулы, помимо завидной подвижности, во многих случаях развили способность к яйцеживорождению и живорождению с подобием плацентарного вскармливания эмбрионов (как у холодолюбивых млекопитающих). Это является эволюционным признаком мобильности, поскольку потомство выводится не из стационарно отложенной икры, а на ходу, что повышает его шансы на распространение по экосреде. Добавим, что и яйцекладущие акулы по-своему мобильны. Они откладывают яйца (икринки) в твердой оболочке, чем уподобляются, например, рептилиям, у первых представителей которых способность откладывать яйца в твердой скорлупе послужила одной из основ сухопутной подвижности.
Лучеперые рыбы и акулы не успели продвинуться в ордовике так сильно, как эндоцератиты, а потому благополучно перенесли позднеордовикское вымирание, более суровое к остальной биоте [184; 239; 836]. Объяснение парадоксального уклонения лучеперых рыб и акул от позднеордовикского вымирания благодаря отставанию от эндоцератитов мы приведем ниже (вместе с рядом аналогичных случаев).
Силурийский (438–408 млн лет назад) и девонский (408–354 млн лет назад) периоды были временем сближения материков и, соответственно, эпохой прибывающего и отступающего потепления. По мере его углубления биопродуктивность среды возрастала, и суша превращалась в притягательную эконишу для консументов, потребителей органики, благо сухопутная флора к тому времени процветала уже не один миллион лет.
В верхнем ордовике под влиянием отступления оледенения сушу освоили норные животные, в которых подозревают членистоногих многоножек [643]. Затем, в позднем силуре (век пржидоли, 414 млн лет назад), землю завоевали такие членистоногие, как паукообразные и непосредственно зафиксированные многоножки [440]. Позже, в нижнем девоне (век зиген, 401–394 млн лет назад), их состав пополнили насекомые [307; 483], чьи крылья размером до 2 см поначалу служили целям терморегуляции, а с повышением размера до 4 см стали применяться для полета [502]. Поскольку полет увеличивал мобильность насекомых, их крылья должны были служить органами движения в некий холодный период. Однако данных на этот счет пока нет.
Обзаведясь фауной беспозвоночных, суша привлекла позвоночных хищников, которые прибыли в новую для себя экосреду в раннем верхнем девоне (век верхний фран, ок. 370 млн лет назад) [128а; 129, с. 420; 130, с. 508]; а также [246; 253; 274]. Вышедшие из пресноводных водоемов [834], первые земноводные были шестипалыми [345]. Они происходили от кистеперых рыб [334], а те – от рыб вроде Styloichthys changae (век поздний лочлов (жедин), нижний девон Китая), связывающих предков бесчелюстных рыбообразных с предками четвероногих позвоночных (земноводных, пресмыкающихся и млекопитающих) [835]. Рожденные межледниковьем, земноводные были маломобильными существами, а потому держались водоемов, привязанные к ним способом размножения (когда икра откладывается в воду, и там же развивается молодь, как у современных лягушек). Можно было бы предсказать, что, попади земноводные под давление отбора на высокомобильность, они изменили бы способ размножения на независимый от водной среды, принявшись прочесывать сушу и обзаведясь для этого непроницаемыми покровами тела взамен слизистой голой кожи, служащей для дыхания. Все это осуществили пресмыкающиеся.
По мере сближения материков климат девона портился, биопродуктивность среды падала, и маломобильные организмы погружались в кризис, что выразилось в позднедевонском вымирании (рубеж веков фран/ фамен, 367 млн лет назад), которое, помимо всего прочего, вызвало гибель тропических коралловых рифов и убыль чистой биомассы [184; 239]. Это прямо отражало снижение палеотемператур (чего не переносили прикрепленные маломобильные коралловые полипы) и оскудение среды.
Вслед за теплым девоном наступили каменноугольный (карбон, 354–286 млн лет назад) и пермский (286–251 млн лет назад) периоды. В это время континенты все более сближались, чтобы образовать сверхматерик Пангея (ок. 250–180 млн лет назад). Эти события вызвали пермо-карбоновое, или гондванское (по южной половине Пангеи – Гондване), оледенение (310–260 млн лет назад). Под давлением отбора на мобильность в соответствии с вышеописанным сценарием земноводные превратились в амниот (от греч. 'αμνίον – околозародышевый пузырь), т. е. начали откладывать яйца в плотной зародышевой оболочке, позволяющей им развиваться на суше. Защищенная чешуей кожа открывала недавним амфибиям сухопутные просторы. Появились пресмыкающиеся (век бригант (визей), нижний карбон, ок. 338 млн лет назад) [716].
На протяжении пермо-карбонового оледенения пресмыкающиеся сохраняли четвероногость, которая обеспечивала им максимальную мобильность. Ошибочно думать, будто ранние пресмыкающиеся не были в состоянии усвоить какую-либо другую локомоцию. Напротив, в их среде появилась интересная ящерицеобразная рептилия Eudibamus cursoris (из числа Bolosauridae, ранняя нижняя пермь, век ассель, 290 млн лет назад) [187]. Она напоминала ряд современных ящериц малопродуктивных, открытых пространств, бегающих рысью, переходящей в бег на двух задних ногах (такова, например, австралийская плащеносная ящерица), что отличается от энергосберегающей шагающей двуногой локомоции динозавро-человеческого типа. Отсюда следует, что E. сursoris был порожден оледенением и не имел ничего общего с двуногими динозаврами и людьми (в смыле конвергентной, внешне похожей эволюции), но подтверждал способность ранних рептилий разнообразить свою походку.
Здесь необходимо остановиться на способах локомоции (передвижения), присущих высшим наземным позвоночным. За вычетом промежуточных вариантов вроде бега E. cursoris или продвинутых кроличье-лягушачьих прыжков кенгуру, тушканчиков и им подобных животных существуют два основных типа локомоции: четвероногая, как у подавляющего большинства современных млекопитающих и «холодолюбивых» динозавров, и шагающая двуногая, как у «теплолюбивых» динозавров, людей и некоторых других существ.
Четвероногость весьма расточительна энергетически, сопряжена со значительным сцеплением животного с субстратом (из-за четырех конечностей), однако обеспечивает ему исключительную выносливость, проходимость, значительные скорости и дистанции передвижения. Подобная адаптация крайне полезна в прохладных условиях низкой биопродуктивности среды, когда скудную пищу приходится собирать с больших пространств.
В противоположность этому шагающая двуногость весьма выгодна, экономна энергетически, не сопряжена с большим сцеплением тела с субстратом (благодаря опоре на две конечности), однако не дает своему обладателю выносливости и не позволяет осваивать значительные территории. Такой способ передвижения выгоден в теплые эпохи высокой биопродуктивности среды, когда пища богата, скучена в пространстве и не требует больших путешествий для своей добычи. При этом двуногое существо очень выигрывает, экономя энергию на передвижении. С конкретными примерами такого рода мы познакомимся ниже.
Пермокарбоновое оледенение отмечено еще двумя перспективными достижениями биосферы. По молекулярно-генетическим данным, в это время возникли цветковые (покрытосеменные) растения. В конце нижнего карбона (век намюр, 319 ± 35 млн лет назад) они разделились на однодольные и двудольные, а в перми (276 ± 33 млн лет назад) двудольные растения претерпели радиацию [545]. Покрытосеменные растения были родственны растениям из рода Glossopteris, преобладавшему в перми и отличавшемуся простым способом размножения, как у цикадовых и гинкго [595]. Происходя из холодного ордовика, все наземные растения имели мобильные средства размножения (споры, семена) и совершенствовались в этой части. Возникнув в холодном карбоне, цветковые растения сделали здесь очередной шаг вперед: благодаря различным приспособлениям их семена распространялись на большие расстояния лучше, чем у голосеменных (например хвойных), хотя последние уступили преобладающее положение не сразу [88, с. 21–382].
Другое достижение касается холодолюбивой фауны, в рамках которой пресмыкающиеся котилозавры породили в верхнем карбоне звероподобных рептилий, господствовавших в перми, постепенно убывающих в триасе, вымирающих в средней юре, однако отмеченных в поздней юре и даже в позднем палеоцене (век ранний тиффан, ок. 60–57,3 млн лет назад) [25, с. 110–117; 87, с. 230–298; 250; 328; 655, с. 173–186]. От этих четвероногих рептилий в перми произошли зверозубые пресмыкающиеся – чрезвычайно прогрессивные существа: некоторые, по-видимому, стали теплокровными, покрылись шерстью и усвоили разнозубость. Зубы прочих пресмыкающихся обычно однородны (за вычетом динозавра гетеродонтозавра из позднего триаса или ранней юры (ок. 201 млн лет назад), у которого самцы имели клыки [37, с. 46–47]; гетеродонтность динозавра Eoraptor lunensis из верхнего триаса (ок. 230 млн лет назад) далека от разнообразия зверозубых рептилий и млекопитающих, cp. [689, с. 64, рис. 1]). У продвинутых же зверозубых пресмыкающихся зубы дифференцированы на резцы, клыки и коренные (как у их потомков – млекопитающих). Это требовалось для эффективного использования небогатой и разреженной пищи ледниковья. Казалось бы, перед прогрессивными зверозубыми рептилиями открыт весь мир. Между тем, их постепенно смело с эволюционной сцены.
С концом пермского похолодания грянуло позднепермское вымирание, неблагосклонное к мобильным организмам. Однако оно лишь потеснило цветковые растения и зверозубых рептилий. Это вымирание осуществлялось в две стадии: гваделупа, ок. 256 млн лет назад, истребившая 58% родов морских организмов, и татарий, ок. 251 млн лет назад, уничтоживший 67% родов морских организмов [723; 825 a]; cp. [35, с. 87, рис. 2; 184; 239; 311; 471; 639; 674, с. 437–439; 802].
Сухопутная биота понесла сходные потери, отвечающие эволюционно-экологической направленности естественного отбора, связанного с потеплением. Закономерно были утрачены небольшие подвижные рыбоеды и насекомоядные существа. Их участь разделили средние и крупные травоядные, привыкшие собирать скудные корма с обширных пространств. Высшие хищники потянулись следом. Даже спустя 15 млн лет этот урон среди пресмыкающихся еще не восполнился [185].
Легко понять, что избирательное двухступенчатое пермо-триасовое вымирание подчинялось определенной эволюционной логике, а не уничтожало живых существ без разбора, как свойственно стихийным бедствиям вроде падений астероидов или извержений вулканов. Как и в случае с мелтретичным вымиранием (см. далее), углубление исследований рисует эти биокатастрофы все менее окказионально (т. е. все менее случайно).
Мезозой (251-65 млн лет назад)
Наступил триасовый период (251–201 млн лет назад), начало которого ознаменовалось «метановым потеплением» (вследствие парникового эффекта от вулканических газов, включая метан) [742] и морской трансгрессией (подъемом уровня Мирового океана) [74, с. 219, рис. 7.1; 401]. Естественный отбор принялся поощрять маломобильных животных. Еще в верхней перми от рептилий-эозухий, близких прогрессивным котилозаврам-капторинам и низшим зверообразным пеликозаврам (заложившим черты молекулярно-генетического родства своих потомков-млекопитающих и людей с потомками текодонтов, т. е. птицами), произошли пресмыкающиеся текодонты (псевдозухии). Под влиянием потепления и связанного с ним роста биопродуктивности среды, они усвоили экономичную шагающую двуногость человеческого типа, длительное время процветали, однако не перенесли похолодания (падения биопродуктивности среды) в конце триасового периода и вымерли. Однако до того, в середине триаса, морская трансгрессия и потепление обострили давление отбора в пользу маломобильных животных. В результате от текодонтов монофилетично (из одного корня) произошли двуногие динозавры двух отрядов (ящеротазовые и птицетазовые), 227,8 ± 0,3 млн лет назад (см. рис. 3) [87, с. 494, 525; 271; 324; 329; 653; 655, с. 138–141; 688–690].
Судя по своим поздним потомкам мелового периода и подобно ближайшим родичам, птицам и крокодилам, динозавры имели четырехкамерное сердце. Аналогично птицам они были покрыты перьями [148; 441; 747; 827–829], возможно, отличались постоянной температурой тела (гомеотермией) [173]; cp. [660] и даже спали в птичьей позе [826]. Они были сходны с птицами в особенностях роста челюсти [172], но расходились с ними в скоростях общего роста [310], что можно объяснить повышенным метаболизмом (обменом веществ) у последних вследствие летающего образа жизни. Кроме того, двуногим динозаврам, даже их верхнемеловому венцу, хищному колоссу тираннозавру, были присущи маломобильность [430] и равнозубость, ориентированная на расточительное потребление богатой и скученной пищи, – то и другое гармонировало с высокой биопродуктивностью среды.
Длительное триасовое потепление раздвинуло существующие экониши и вовлекло в них маломобильных существ, что привело пресмыкающихся в водную среду и дало известных ихтиозавров (средний триас – поздний мел), плезиозавров (тогда же) и других водных ящеров [25, с. 18, 22, 33]. Похолодание позднего триаса (норий-рэтийская морская регрессия, падение уровня Мирового океана) привело к упадку и частичному вымиранию маломобильных ихтиозавров. Выжили те их формы, которые лучше освоили живорождение, являющееся вариантом неявной мобильности.
То же похолодание отпочковало от динозавров подвижных летающих птиц, засвидетельствованных спорными остатками Protoavis texensis («первоптицы из Техаса», поздний триас, ок. 225 млн лет назад [141]; cp. [240; 326]), ископаемыми птичьими следами (поздний триас, века норий-рэт, св. 212,5 ± 7 млн лет назад [565]) и синхронным молекулярно-генетическим расхождением птиц с крокодилами (222 ± 52 млн лет назад [479]), что, впрочем, не исключает обособления предков птиц от крокодилов еще в составе динозавров (т. е. сначала от крокодилов отделились динозавры, а впоследствии, много позже, от динозавров обособились птицы вроде археоптерикса, которые положили начало современной авиафауне, что представляется нам сомнительным, см. ниже).

Рис. 3. Ставрикозавр (Staurikosaurus pricei, «ящер созвездия Южного Креста») из семейства ставрикозаврид подотряда теропод (звероногов) отряда заурисхий (ящеротазовых динозавров) был ловким двуногим хищником и охотился в среднем триасе, не подозревая, что в конце этого периода его отсталые родичи прозауроподы встанут на четвереньки и дадут начало крупнейшим наземным животным планеты, зауроподам. [37, с. 30–31]
Позднетриасовое вымирание (210 млн лет назад [184; 239; 518]) затронуло малоподвижных животных, но оживило четвероногих, что выразилось в оформлении млекопитающих как класса позвоночных животных (век норий, св. 200 млн лет назад [330]). Однако затем рэтийская регрессия ненадолго сменилась парниковым потеплением на 3–4° вследствие роста вулканического диоксида углерода 205,7 ± 4 млн лет назад [560], что спровоцировало вымирание мегафлоры, ориентированной на падение биопродуктивности среды.
Наступивший юрский период (201–144,2 млн лет назад) открылся очередной регрессией (похолоданием) и потряс двуногих динозавров – произошло их ограниченное нижнеюрское вымирание [239, с. 49]. Если целурозавры и карнозавры всегда оставались удачливыми, а потому эволюционно негибкими двуногами, то неуклюжие двуногие прозауроподы были готовы ко всему новому (о причинах чего см. далее) и охотно перешли к четвероногой локомоции, превратившись в гигантских зауропод (нижняя юра, век поздний лейас – верхний мел) (см. рис. 4, 5).
Отметим впечатляющую парадоксальность эволюционного процесса: предки динозавров (текодонты) сначала «встают» с четырех ног на две, порождают успешных двуногих динозавров, а затем вновь «опускаются» на четыре конечности (как произошло в случае с зауроподами и некоторыми другими динозаврами, см. далее). В рамках нашей версии эволюционной направленности этот нонсенс находит естественное объяснение – климато-экологическое. Возможно, обсуждаемое нижнеюрское похолодание развило у гетеродонтозавров разнозубость, являющуюся адаптацией к низкой биопродуктивности среды, свойственной похолоданиям.
После метанового потепления (ок.183 млн лет назад [742]) в середине средней юры (век поздний байос) палеотемпературы упали с наступлением антарктического микрооледенения, о чем свидетельствуют тиллиты (ископаемые морены, ледниковые отложения) юрского времени в Антарктиде [42, с. 33; 74, с. 220, 222], однако затем тепло стало нарастать к максимуму трансгрессии в середине верхней юры (конец века кимериджа). Средне-верхнеюрское потепление пагубно сказалось на четвероногих звероподобных рептилиях, которые почти полностью вымерли, оставив по себе молодых отпрысков, млекопитающих. То же самое произошло с четвероногими зауроподами, испытавшими с потеплением первый эпизод вымирания [25, с. 56]. Их судьбу повторили четвероногие птицетазовые динозавры сцелидозавры из ранней юры, когда их четвероногость была закономерна. Во второй половине юрского периода они дали четвероногих же стегозавров (середина юры – ранний мел), а через сирмозавров (конец раннего мела) – панцирных анкилозавров (ранний – поздний мел) [25, с. 79]. Иными словами, эволюция этих существ шла как бы «ракетным способом», когда в моменты потеплений отбрасываются «отработанные» (устаревшие и вымирающие) ступени.
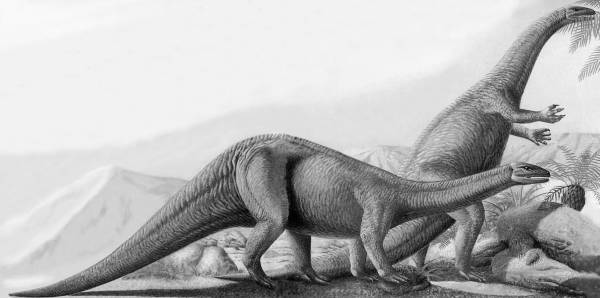
Рис. 4. Платеозавр (Plateosaurus engelhardti, плоский ящер Энгельгардта) из семейства платеозаврид инфраотряда прозауропод (проящероногов) из подотряда теропод группы зауроподоморфов (ящероногоподобных) отряда заурисхий. Это неуклюжее двуногое животное питалось растительностью, но происходило от совершенных двуногих динозавров среднего триасового периода, хищных теропод. В позднем триасе под влиянием похолодания и падения биопродуктивности среды прозауроподы стали возвращаться к четвероногому способу передвижения, свойственному далеким «холодолюбивым» предкам двуногих текодонтов и динозавров [37, с. 34–35]
Нижнеюрское похолодание с его давлением в пользу мобильности выщепило из среды архозавров (родственников динозавров) летающих ящеров птерозавров, покрытых шерстью. Они просуществовали до верхнего мела и разделили судьбу динозавровой биоты [25, с. 106–109]. Их вымирание объясняется тем, что, в отличие от сноровистых, высокоподвижных птиц современного типа, они были планирующими, энергосберегающими созданиями, а не активными летунами, т. е. характеризовались ограниченной мобильностью [25, с. 106; 87, с. 593], в то время как маастрихтское (конца мела) и палеоценовое (после мела) похолодания требовали от животных более сильно выраженной подвижности.

Рис. 5. Барапазавр (Barapasaurus tagorsi, большеногий ящер) из подотряда зауропод (ящероногов) группы зауроподоморфов отряда заурисхий. Под влиянием ухудшения климата и падения биопродуктивности среды неуклюжие двуногие прозауроподы породили в раннем юрском периоде совершенных четвероногих зауропод. Они процветали и достигли размеров, рекордных для наземных животных [37, с. 42–43]
В поздней юре (150,7–144,2 млн лет назад) максимум трансгрессии сменился регрессией и похолоданием, что возобновило селекцию на подвижность и побудило динозавров вторично породить птицеподобных существ – в частности, известного археоптерикса [240, с. 349], который строением зрительной и слуховой коры головного мозга действительно напоминал современных птиц [136], что явилось результатом конвергенции, параллельной эволюции. Будучи предком энантиорнитинов, или «противоположных птиц» (от последовательности совмещения костей стопы в ходе развития, противоположной современным птицам), археоптерикс напоминал миниатюрного велоцираптора или дромеозавра [697], тоже пернатых динозавров, и отличался от протоависа, предка орнитуринов, давших современных птиц. Динозавров, археоптерикса и энантиорнитинов объединяли примитивные признаки эктотермности, т. е. непостоянной температуры тела [660].
Меловой период (144,2–65 млн лет назад) ознаменовался нижнемеловой регрессией (век валанжин, 130 млн лет назад) и продолжением отбора на мобильность. В это время развивались упомянутые тупиковые энантиорнитины и перспективные орнитурины, в среде которых началась дивергенция современных птиц (средний нижний мел, век апт, 122,9 ± 0,3, 121,2 ± 0,3–121,1 ± 0,2 млн лет назад) [260; 747]. За тот же временной интервал пернатые динозавры породили еще одно птицеобразное создание – четырехкрылого Microraptor gui из Китая [829], все четыре конечности которого годились для планирования, повышая подвижность. Среди современников этого маленького дромеозавра – китайских птиц-энантиорнитинов – также встречался четырехкрылый вид [833], так что данная эволюционная тенденция проявлялась неоднократно.
Если зауроподы откликнулись на падение биопродуктивности среды мобильной четвероногостью, то орнитоподы, ящеротазовые четвероноги (игуанодоны) и двуноги (утконосые динозавры-гадрозавры, верхняя юра – верхний мел) отреагировали иначе: они освоили умение жевать, как млекопитающие [596], что повышало эффективность потребления редкой пищи соответственно похолоданию. В этом отношении названные представители динозаврового племени принципиально отличались от прочих архозавров, т. е. других динозавров, а также крокодилов. Последние, кстати, пережили роковой меловой период в силу того обстоятельства, что являлись ловкими наземными бегунами-четвероногами, т. е. мобильными животными, что открывало им путь в холодный кайнозой, где они, впрочем, вскоре «замерзли» и перешли к водному образу жизни в тропических и субтропических реках, не выдержав конкуренции с крепнущими наземными четвероногими млекопитающими.
Рубеж нижнего-верхнего мела (105 млн лет назад) прошел с трансгрессией, а века турон – коньяк (92–86 млн лет назад) подверглись вулканическому диоксид-углеродному потеплению [750]. В конце мела, в маастрихте (71–66 млн лет назад) регрессия сопровождалась заметным похолоданием [526], когда тропические температуры в океанах упали ниже современных [288]. К этому времени изнеженные теплым и обильным мезозоем (251–65 млн лет назад) динозавры находились на вершине своего расцвета, о чем свидетельствует сам факт существования крупнейшего наземного хищника – королевского тираннозавра (76–65 млн лет назад) (см. рис. 6). Сопряженная с ним мегафауна стала совершенной и высокоспециализированной, а потому могла лишь углубляться в аппетитах, но никак не ограничивать себя, чего требовала оскудевающая обстановка.
Вымирание динозавров происходило постепенно и пропорционально прогрессу млекопитающих (по модели цветковых растений и костистых рыб, см. далее). В веке кампан (76–73 млн лет назад) на пик расцвета динозавров приходилось 30 их родов. С приближением маастрихтского похолодания (71–66 млн лет назад) гордые животные ощутили эволюционно-экологическое затруднение: они любили много потреблять и мало двигаться, а теперь среда благоволила к сноровистым аскетам типа четвероногих млекопитающих.

Рис. 6. Тираннозавр (Tyrannosaurus rex, королевский ящер-тиран) из семейства тираннозаврид инфраотряда карнозавров (хищных ящеров) подотряда теропод отряда заурисхий. Это был крупнейший наземный хищник всех времен, чуть уступавший лишь хищному гигантозавру начала верхнего мела (ок. 100 млн лет назад), который, будучи южноамериканцем, закономерно превосходил массой североамериканского собрата, как свойственно рептилиям с приближением к экватору (у млекопитающих – наоборот). Тираннозавр был совершенным хищником (и падальщиком, как современные львы), узко приспособленным к среде, богатой пищей. С ухудшением климата и падением биопродуктивности среды королевский ящер угас и вымер вместе с подавляющим большинством остальных динозавров. По современным данным, он был оперенным существом [37, с. 98–99]
За весь маастрихт и две трети дания (72–65,3 млн лет назад) число родов динозавров упало до 12. Затем вымирание ускорилось, и за период 65,3–64, 96 млн лет назад прошло пять убывающих стадий: 12–12–11–10–7 родов уходящих динозавров. О том, что биосфера отреагировала на коварный удар иридиевого астероида 65 млн лет назад, не может быть и речи. Параллельно со ступенчатым падением динозавров шел уверенный прогресс копытных млекопитающих, представленных в тех же временных точках 0–1 – 3–5 – 8 родами. Так что за 40 тыс. лет, истекших с момента падения астероида, млекопитающие зримо окрепли, а динозавры еще не вымерли окончательно, являя редкий случай послемеловых, палеоценовых динозавров, останки которых обнаружены выше мелтретичной иридиевой аномалии, оставленной упавшим болидом [714; 713]; cp. [222; 642; 696].
Приведенные количественные данные по динозаврам и копытным млекопитающим за период 65,3–64,96 млн лет назад происходят из формации Хелл Крик в Монтане, США (северо-запад страны). Монтана лежит на том же континенте, что и мексиканский кратер Чикхулуб, оставленный ударным небесным телом 65 млн лет назад. Монтанская биота совершенно не отреагировала на это событие: млекопитающие набирали эволюционные обороты, а динозавры сокращались в разнообразии в соответствии с тенденцией, наметившейся за 7 млн лет до вторжения астероида. Не избежали их судьбы и обитатели других сред.
Морские пресмыкающиеся (ихтиозавры, плезиозавры, мозозавры и пр.), страдавшие низкой мобильностью [35, с. 205–206], вымерли в верхнем меле: более подвижные ихтиозавры – в его теплом начале, остальные – ближе к холодному концу [25, с. 18, 33]. Господствовавших на море раковинных головоногих моллюсков аммонитов и белемнитов постигла та же участь.
Показательна в этой связи эволюционная история теплолюбивых аммоноидей (нижний девон – верхний мел) [83, с. 243–425; 84, 15–144]. Появившись в начале раннего девона (век жедин, 408–401 млн лет назад), они продолжили линию других головоногих моллюсков – бактритов, имевших прямую или слабосогнутую раковину, т. е. являвшихся более-менее хорошими пловцами. Однако «возниковение аммоноидей было связано с образованием вначале согнутой, а затем криоконовой раковины, состоявшей из несоприкасающихся оборотов, свернутых в одной плоскости» [38, с. 376]. Это делало аммоноидей слабыми пловцами и приводило в соответствие с теплым, высокобиопродуктивным девоном, когда была выгодна умеренная мобильность.
В эпоху пермо-карбонового оледенения ситуация изменилась, и малоподвижные аммоноидеи закономерно отступили в эволюционную тень, поскольку оледенение благоприятствовало более энергичным созданиям. Зато теплый мезозой востребовал их качества, и в кампане – маастрихте (76–66 млн лет назад) они выдвинули род Pachydiscus, у представителей которого свернутая раковина достигала рекордных 2 м в диаметре [84, с. 111]. Напомним, что скоростное плавание обеспечивала прямая раковина (см. выше об эндоцератитах). Свернутая же в диск раковина, наоборот, скоростей не давала. При этом под влиянием климатических колебаний в меле аммониты то пытались слепо распрямить раковину, чтобы повысить свою мобильность и попасть в унисон с похолоданиями, то сворачивали ее обратно, чтобы убавить подвижность и совпасть с потеплениями. В силу закона Долло о необратимости эволюции эти попытки были обречены на провал и лишь наводняли меловые моря уродливыми созданиями.
Избирательность верхнемелового вымирания прослеживается в судьбе двустворчатых моллюсков. Их прикрепленные формы вымирали заметно интенсивнее, нежели свободноживущие [434]. Это объясняется тем, что прохладное дыхание наступающего кайнозоя готовило перспективу лишь подвижным существам – в данном случае свободноживущим двустворчатым моллюскам.
С мезозойскими воздухоплавателями происходило то же самое. Отряды подвижных современных птиц дивергировали еще при похолодании в раннем меле, и 22 их филетические линии успешно пересекли мелтретичную границу, не побоявшись зловещего астероида [260]; ср. [252]. Однако сам факт биологической принадлежности к современным птицам отнюдь ничего не значил. В частности, могучие (более метра в высоту) нелетающие гесперорнисы были водоплавающими птицами и в силу своей маломобильности не перенесли верхнемелового отбора. Маломобильным планирующим птерозаврам тоже не повезло, равно как и «противоположным птицам» энантиорнитинам, которые летали хуже тогдашних орнитуринов, предков неорнитов, нынешних птиц [697].
Между тем, финал мелового периода не был эпохой тотального вымирания [35, с. 89, рис. 3; 184; 239; 394; 407; 674, с. 439–441]; cp. [444]. Эволюционная история меловой биоты выглядела как пара сообщающихся сосудов, в одной из емкостей которых убывали многочисленные маломобильные господа мезозоя (динозавры, птерозавры, ихтиозавры, плезиозавры, аммониты, белемниты и др.), а в другой прибывали скромные высокомобильные господа будущего кайнозоя (покрытосеменные растения, млекопитающие, современные птицы, костистые рыбы, акулы, внутреннераковинные головоногие моллюски и др.).
Будучи родом из пермо-карбонового оледенения (см. выше, а также [284; 407; 638]), цветковые растения положительно отреагировали на ухудшающуюся климатическую обстановку нижнего мела и в середине мела (кон. века альба, 105 млн лет назад) произвели революцию флоры. С верхнего мела к ним перешло флористическое господство, которое укрепилось в кайнозое и наиболее ярко представлено сейчас [88, с. 423].
Через своих рептильных звероподобных предков млекопитающие тоже ведут начало из пермо-карбонового оледенения. Оттуда происходит их четвероногая мобильность и эффективная разнозубость, выгодные в условиях скудной биопродуктивности среды. Как класс, они выщепились в холодное время норий-рэтийской регрессии. В поздней юре или раннем меле из их пула выделились однопроходные звери (ок. 144,2 млн лет назад) [149; 466]. По молекулярно-генетическим данным, то же самое произошло с сумчатыми млекопитающими еще раньше (в средней юре, 173 ± 12,3 млн лет назад) [479], хотя палеонтологически они известны лишь с позднего мела (84–65 млн лет назад) [183]. По молекулярно-генетическим данным, пять линий высших плацентарных млекопитающих обособились св. 100 млн лет назад, т. е. в начале верхнего мела (в веке сеномане, 105–92 млн лет назад), и большинство их современных отрядов появилось до мелтретичной границы; палеонтологически плацентарные млекопитающие отмечены 85 млн лет назад [479]; а также [651; 655, с. 187–310]. Радиация сумчатых и плацентарных млекопитающих связана с теплыми периодами юры и мела, когда подавляющие их четвероногие динозавры испытывали эволюционную депрессию, и вакансии в соответствующих эконишах доставались млекопитающим. Последние были преимущественно мелкими формами с черно-белым зрением, поскольку вели скрытный, ночной образ жизни. Однако среди них встречались и более выразительные представители. В раннем меле Китая, 139–128 млн лет назад, существовал хищник из числа млекопитающих Repenomomus giganticus, достигавший свыше метра в длину и 12–14 кг массы, что позволяло ему успешно охотиться на детенышей динозавров (в частности, на пситтакозавров, один из которых сохранился у него в брюхе [425]). Млекопитающие, подвижные зверьки, настойчиво прогрессировали и, подобно птицам, уверенно пересекли мел-палеоценовую границу.
Параллельно цветковым растениям, млекопитающим и птицам меловую экосреду завоевывали костистые рыбы. Являясь отпрысками лучеперых рыб, предки которых зародились в период ордовикского оледенения, костистые пловцы были сильны в прохладной, малопродуктивной среде, поскольку лучше других рыбообразных создавали осевым скелетом двигательную локомоторную волну [124]. Недаром их современный рекордсмен, меч-рыба, развивает огромную скорость до 130 км/ч, превосходя по этой части наземного гепарда. Непреодолимо прогрессируя с раннемелового похолодания, они возобладали в морях с верхнего мела, легко пересекли мелтретичную границу и ныне составляют 95% ихтиофауны [86, с. 334; 655, с. 61–71]. Эта победа облегчалась малой мобильностью прочих мезозойских рыб [35, с. 205]. Эволюционная история акул, также имеющих ордовикские корни и отличающихся высокими ходовыми качествами, развивалась по образцу костистых рыб [86, с. 196–237] и тоже отмечена рекордами в кайнозое (см. ниже). Наконец, безуспешная борьба за жизнь медлительных аммонитов и белемнитов закончилась вытеснением их подвижными внутреннераковинными головоногими моллюсками: десятиногими кальмарами и каракатицами, а также и восьминогими осьминогами [35, с. 187; 84, с. 145–178].
С падением общеглобальной биопродуктивности среды разнообразие жизни на планете убывает, и проявления эволюционной направленности теряют разнообразие. Масштабы организмов тоже мельчают. Поэтому господствующие наземные животные мезозоя были крупнее кайнозойских, а раннекайнозойские – внушительнее позднекайнозойских современников нарастающего похолодания. Соответственно, примеры нашей версии эволюции для кайнозоя относительно скромны.
Кайнозой (65-0 млн лет назад)
В палеоцене (65–55 млн лет назад) высокомобильные организмы мелового периода продолжают разнообразиться, не меняя своей эволюционной направленности, поскольку вся эпоха отмечена регрессией и похолоданием. В начале эоцена (55–36 млн лет назад) началось метановое потепление [742], и регрессия сменилась трансгрессией. Следовало бы ожидать, что в установившихся теплых условиях появятся маломобильные формы мезозойского типа.
В соответствии с этим ожиданием в среднем эоцене распространяются уцелевшие потомки вымерших меловых белемнитов – байанотеутиды [35, с. 189–190; 38, с. 382, 388, рис. 217; 84, с. 161–162]. Тогда же возникают не связанные общностью происхождения нелетающие, бегающие птицы Ratidae, своего рода конвергенты (т. е. внешние подобия) двуногих динозавров: страусоподобные элеутерорнисы (средний эоцен, век лютет) и эпиорнисовые псамморнисы (эоцен) [35, с. 245–246]. Наконец, в раннем среднем эоцене Германии (50–48 млн лет назад) обнаруживается двуногое бегающее млекопитающее Leptictidium nasutum, подлинный конвергент двуногих динозавров со свойственным им и людям прямохождением (см. рис. 7) [118, с. 34]. Эоценовое потепление повысило биопродуктивность среды и раздвинуло рамки экониш, привлекая в них новых обитателей. В соответствии с этой новацией сухопутные парнокопытные произвели водных китообразных в нижне-среднем эоцене (ок. 52 млн лет назад [752]) или в конце среднего эоцена (в лютете, 47 млн лет назад [354]). В гармонии со своим происхождением в теплую эпоху китообразные (зубатые киты) развили чуждую млекопитающим равнозубость динозаврового типа.
В олигоцене (36–24 млн лет назад) трансгрессия прекращается и сменяется регрессией во второй половине эпохи. В связи с похолоданием высокомобильные четвероногие млекопитающие испытывают эволюционный подъем и порождают мегафауну, украшением которой является гигантский безрогий носорог Indricotherium transouralicum – самое крупное и высокое наземное млекопитающее, до 5 м высотой (средний олигоцен).

Рис. 7. Легкобег носатый (Leptictidium nasutum) возник в эпоху раннего среднего эоцена на островах нынешней Германии 50–48 млн лет назад, когда из-за потепления поднялась биопродуктивность среды, и естественный отбор дал шанс появлению маломобильных двуногих животных. «Носач» бегал на двух ногах, как двуногие динозавры и люди, а потому являлся их эволюционным конвергентом (подобием), таким же «теплолюбивым», как и они [118, с. 34]
В миоцене (24–5,3 млн лет назад) ранне-среднемиоценовое потепление (24–13 млн лет назад) положило конец олигоценовой мегафауне. Зато пробившийся отбор на маломобильность вызвал к жизни двуногую обезьяну «перелоапитека каталонского» (Испания, 13 млн лет назад), положившего начало целой плеяде двуногих гоминин (см. разд. 2. 4). Происходя от аборигенов высокобиопродуктивных тропических лесов, они органично несли в себе мезозойскую ориентацию на маломобильность и прожорливость, что в сочетании с эволюционной отсталостью создало гоминин, перспективных господ будущего, ставших лидерами именно в силу своей отсталости.
В плиоцене (5,3–1,6 млн лет назад) начало эпохи ознаменовалось трансгрессией. Затем последовал ряд гляциалов (оледенений), интергляциалов (межледниковий), стадиалов (пиков оледенений) и интерстадиалов (смягчений оледенений) выраженной ледниковой эпохи. Ее «лицом» можно назвать тридцатиметровую белую акулу Carcharodon megalodon, вымерщую в среднем плиоцене (3,3 млн лет назад), когда интергляциальная трансгрессия Астий вызвала ограниченное вымирание в среде мобильной биоты; cp. [680].
Наконец, в плейстоцене (1,6–0,0117 млн лет назад) развилась богатая приледниковая мамонтовая фауна, вымершая с началом голоценового потепления 11700 календарных лет назад, или 10200 лет назад по радиоуглероду 14С (поясним, что вследствие колебаний содержания тяжелого изотопа углерода 14С в древней атмосфере радиоуглеродные даты исследуемых образцов оказываются моложе своего реального, календарного возраста, который можно установить, например, методом дендрохронологии, когда высчитываются годичные кольца очень старых деревьев, до 8000 лет возраста, после чего итог сравнивается с радиоуглеродной оценкой; получается, например, что календарной дате 4650 лет назад отвечает радиоуглеродная дата 4050 лет назад).
Подобно динозаврам, мамонты и их спутники вымерли не сразу. Некоторые виды жили дольше: островные карликовые мамонты острова Сен-Пол в Беринговом море, 7908 ± 100 лет назад [391], островов Санта-Барбара в Калифорнии [480, с. 213], острова Врангеля на границе Восточно-Сибирского и Чукотского морей, 7620–3730 лет назад [515; 768]; большерогий олень в Западной Сибири, 6900 14C, или 7700 календарных лет назад [736]; возможно, шерстистый носорог в Чувашии, 921/922 гг. н. э. [54, с. 61–66, 139–140; 91, с. 72–74].
«Лицом» нынешней голоценовой эпохи – межледниковой, однако все еще не свободной ото льдов – можно считать гигантского кальмара Architeuthis longimana 19 м длиной (возможны тридцатиметровые особи), продолжающего линию внутреннераковинных головоногих моллюсков, которые сменили аммонитов в конце мелового периода.
Относительно упомянутых вымираний существуют неокатастрофические взгляды, касающиеся в первую очередь пермо-триасовых и мел-третичных событий. Согласно одному из них, массовые вымирания вызваны вулканической деятельностью планеты, которая обескислороживает океаны, отравляя их. Эти несчастья происходят с периодичностью в ~ 32 млн лет [632]. Они ответственны за массовую гибель биосферы на границе перми и триаса [227; 637; 639], а также мела и палеоцена [227; 598]; cp. [361, с. 368].
Показано, однако, что обескислороживание океанов господствовало на Земле на протяжении 250 млн лет в позднем неопротерозое ок. 800–543 млн лет назад [471; 802], однако на тогдашней жизни (например на морской эдиакарской фауне) это заметно не сказалось. Показано также, что обескислороживание океанов в самом начале триаса явилось следствием предшествующего позднепермского вымирания (кризиса радиолярий, морских микроорганизмов) [633], а никак не его причиной. Наконец, показано, что обескислороживание океанов имело место и в конце нижнего мела (в альбе, ок. 112 млн лет назад) [481], что не помешало костистым рыбам, акулам и прочим наследникам мезозоя уверенно набирать эволюционные обороты. В свете указанных расхождений с реальностью вулканическая гипотеза не может быть принята.
Согласно другим представлениям, Землю бомбардируют космические тела (астероиды и кометы), удары которых поднимают в атмосферу взрывной материал, затеняют им планету и создают эффект «ядерной зимы», которая умерщвляет биосферу. О падениях небесных объектов говорят геологические прослойки редкого металла иридия, встречающегося в метеоритном веществе, как, впрочем, и в земных породах, изверженных из мантии в результате вулканической деятельности. Последнее обстоятельство заставляет исключить ископаемую иридиевую аномалию, например ордовик-силурийской границы, из числа космогенных [804]. В разведанной истории Земли насчитывают до 140 болидовых ударов [347]; cp. [361, с. 368]. Среди них наиболее известны следующие:
• поздний плиоцен, ок. 2,15 млн лет назад [347] или ок. 2,3 млн лет назад [482] (астероид св. 1 км в поперечнике);
• средний плиоцен, 3,3 млн лет назад [680];
• поздний эоцен, 35,7 ± 0,2 млн лет назад [199];
• ранний эоцен, ок. 50 млн лет назад [347];
• мелтретичный, 65 млн лет назад [137; 138; 192; 195; 378; 415; 433; 461–463; 477; 554; 558; 561; 563; 649; 743; 745]; болид вызвал глобальные лесные пожары [414; 771];
• поздняя юра, 140 ± 20 млн лет назад [378];
• триас-юрский [603];
• поздний триас, 211 (220–200) млн лет назад [518, с. 46]; 5 кратеров, век норий, 214 млн лет назад [721; 786].
• пермо-триасовый [175]; (251,4 млн лет назад) [180]; cp. [311].
• верхний девон, рубеж веков фран-фамен, 367 млн лет назад [247].
• средний девон, ок. 380 млн лет назад [305].
• поздний докембрий, ок. 600 млн лет назад [365; 366; 806].
• ранний протерозой, 1,85 млрд лет назад [179; 588].
• верхний ранний архей, 3,470 ± 0,002 млрд лет назад [226].
В настоящее время доказано, что после триас-юрской иридиевой аномалии, свидетельствующей о падении болида, тогдашние динозавры отнюдь не вымерли, а, наоборот, расцвели менее чем через 10 тыс. лет после грозного события [603]. Выше мы привели факты, указывающие на то, что позднепермское вымирание состояло из двух крупных стадий, разделенных 5 млн лет, так что его космический источник нереален. Приведены также факты, рисующие позднемеловое вымирание как избирательное событие в духе дарвиновской теории естественного отбора – его космический источник излишен. Кроме того, хорошо известно, что с ранней юры до позднего мела холодные высокие широты Земли населялись полярными динозаврами. Они известны из ранней юры (век плинсбах) Антарктиды [397], из раннего мела (века валанжин – альб, 130–105 млн лет назад) тогдашней заполярной Австралии [644; 142], из позднего мела (века поздний кампан – ранний маастрихт, 76–66 млн лет назад) тогдашней заполярной Аляски [215]. Хорошо приспособленные к продолжительному холоду и тьме полярной ночи, они бы встретили «ядерную зиму» с энтузиазмом. Приведенные соображения делают биологическую сторону космической гипотезы неприемлемой.
Наконец, существует убеждение, что массовые вымирания последних 50 тыс. лет вызваны охотничьим нажимом первобытного человека. Так были истреблены гигантские нелетающие птицы моа Новой Зеландии 980 ± 40–900 ± 45 лет назад [421; 480, с. 222], предположительно мамонтовая приледниковая фауна Старого Света и вся мегафауна Нового Света на рубеже плейстоцена – голоцена 11700 календарных лет назад [480, с. 221–222; 735], мегафауна Австралии 50 ± 5 тыс. лет назад в результате вторжения людей 55 ± 5 тыс. лет назад [570].
Доказано, однако, что мегафауна Австралии вымерла ок. 46,4 (51,2–39,8) тыс. лет назад из-за крайней аридности (засушливости), вызванной гляциальным максимумом [647], т. е. холодным стадиалом Вюрм II C (46,5–39 тыс. лет назад), когда вместе с похолоданием резко обострилась сухость земного климата, поскольку масса воды, в том числе атмосферной влаги, была скована в обширных ледниках.
Кроме того, обнаружено, что Новый Свет был заселен ранее 33 тыс. лет назад, о чем свидетельствуют остатки первобытной культуры в Монте Верде (Чили, 33370 ± 530 лет назад) и Тока ду Бокейрану да Педра Фурада (Бразилия, св. 32 тыс. лет назад) [385; 500, с. 708], в силу чего заселение Америки не может быть увязано с вымиранием ее мегафауны 11 700 лет назад.
Указанные обстоятельства не позволяют принять антропогенную гипотезу массовых вымираний, т. е. древних массовых вымираний, вызванных человеком (за вычетом новозеландского эпизода с птицами моа, который, впрочем, не относится к числу примеров массовых вымираний). Следует добавить, что вулканическая, космическая и антропогенная гипотезы массовых вымираний полностью оторваны от закономерностей развития земной жизни и не проливают здесь никакого света. Вдобавок людям, в том числе и ученым, вообще свойственно переоценивать значение катастроф в истории природы и общества и хвататься за них, как за соломинку, когда что-либо не сразу поддается объяснению (к сказанному добавим неокатастрофическое объяснение происхождения самосознания Дж. Джейнсом, см. разд. 3.2). Между тем, реальное значение катастроф не так фатально. Например, катастрофическое извержение вулкана Кракатау (Индонезия, 1883 г., погибло 36 тыс. человек) или удар Тунгусского метеорита (Красноярский край, Россия, 1908 г., произошел вывал леса в радиусе 15–30 км вокруг эпицентра) не оказали на развитие биосферы никакого влияния.
В изложенной нами версии эволюции биосферы под влиянием эколого-климатического воздействия суперконтинентального цикла бросается в глаза то, что его крупномасштабная периодичность не объясняет более дробных колебаний климата. В реальности суперконтинентальный цикл создает лишь общие условия для развития оледенений или межледниковий. Их конкретные проявления подчиняются влиянию частных факторов астрономического, атмосферного, гидросферного характера, хронометраж которых пока не разработан [70, с. 20; 560; 635; 742; 750], если не считать климатических последствий от колебаний в орбитальном движении Земли с периодами в 25, 44, 100, 133 и 400 тыс. лет [602]. Хронометраж заметных вымираний приводит к выводу, что их пики отстоят друг от друга на 20–60 млн лет [184]: на 26 млн лет за последние 150 млн лет [239, с. 50; 361, с. 363; 410] или на 27 млн лет за последние 250 млн лет (8–12 пиков) [634]. Комментарии этих наблюдений пока преждевременны.
Если принять нашу версию направленной эволюции, то встает вопрос, как она согласуется с дарвиновской моделью случайного хода эволюции и каков генетический механизм направленной эволюции. Мы отвечаем на него следующим образом.
Земная биосфера испытывала климато-экологическое давление среды на протяжении более чем 3,85 млрд лет, и это давление как таковое являлось одним из внешних условий, к которым живые существа приспосабливались в результате случайного естественного отбора. Следовательно, их геномы давно должны были «обзавестись» наследственным механизмом ожидания климато-экологических последствий суперконтинентального цикла, т. е. фактически усвоить склонность к эволюционной направленности естественным путем. В сущности, мы остаемся в рамках дарвинизма, хотя это учение избегало представлений об эволюционной направленности.
Геномы организмов слепы: они лишены органов чувств. Между тем, уже в течение нескольких миллиардов лет незрячие геномы с успехом противостоят изменчивой среде, все больше совершенствуются и, похоже, умеют предвидеть изменения в природе. Как же им это удается? Ответ здесь может быть один. Слепые геномы ориентируются в окружающем мире на основе принципов теории игр.
Разумеется, крохотные геномы не сильны в теории. Однако практическая жизнь и безжалостный естественный отбор сохранили в их рядах только те образцы, которые действуют так, словно и впрямь знакомы с Дж. фон Нейманом (1903–1957, основоположником теории игр). Для разъяснения их «талантов» нам не понадобится высшая математика. Допустим, вы втянуты в азартную игру, а глаза у вас завязаны. Как вы построите свое поведение? По-видимому, пойдете с крупной карты. А если не получится, то с мелкой. Если не выйдет, ограничитесь чем-то средним. И будете так экспериментировать, пока не зацепитесь за удачу. Геномы ведут себя точно так же.
Они находятся в состоянии постоянного мутагенеза, т. е. подвергают свои гены тем или иным мутациям, происходящим с примерно одинаковыми скоростями (ср., однако, разд. 2.4). Вопреки бытующему убеждению, результаты этого процесса далеко не произвольны, не случайны. Дело в том, что генные мутации делятся на серии характерных типов: гиперморфные (они увеличивают проявление подчиненного себе признака, фена), гипоморфные (они уменьшают проявление признака), антиморфные (признак уничтожается вообще) и неоморфные (признак дополняется) [7, с. 42–49]. Иными словами, мутагенез носит количественный характер (как и ваше «слепое» поведение в мысленном эксперименте с азартной игрой). Изменения блоков генов и целых хромосом также подчиняются количественному принципу: они дуплицируются, т. е. удваиваются, умножаются, как и отдельные гены [82; 468; 522; 523; 613].
Что такой мутагенез может дать геному и его организму-хозяину? Уверенность в завтрашнем дне. Допустим, грядет похолодание. Организмы слепо, механически мутируют обрисованным количественным способом. Треть из них немотивированно мутирует гипоморфно и утрачивает, например, запасы жира – они замерзнут. Другая треть вообще воздерживается от мутаций и прозябает. Зато оставшаяся треть, не подозревая о своей гениальности, опять же слепо, механически предается гиперморфным мутациям и ни с того ни с сего наращивает подкожный слой жира. Приходит холод, эта треть не мерзнет и овладевает миром. Она побеждает автоматически, в силу законов теории игр. На Земле уцелели лишь те организмы, которые неукоснительно следовали описанному способу количественного мутагенеза. Он позволял слепо предугадывать климато-экологические последствия суперконтинентального цикла и гарантировал жизнь определенной части участников событий. Другая часть так же гарантированно «не попадала» в ход событий и беспомощно вымирала.
При таком положении дел в эволюции мы могли бы ожидать, что определенный процент организмов в истории жизни будет вымирать систематически и внешне беспричинно – в силу того, что эти организмы не угадали правильный ход в мутагенезе. Но так ли это? Это так. Палеонтологи действительно обнаружили в ископаемой летописи загадочную череду автоматически текущих вымираний, которую они окрестили Законом Постоянного Вымирания, или гипотезой Красной Королевы (персонажа из «Алисы в стране чудес» Л. Кэрролла) [182]. В свете вышеизложенного мы начинаем понимать, откуда в палеонтологической летописи взялся след Красной Королевы.
Теперь посмотрим, что дает версия количественного мутагенеза для понимания узловых моментов истории жизни. Можно было бы ждать, что организмы, угадавшие грядущие перемены в экосреде, окажутся приспособленными к ней наиболее точно и займут в ее ландшафте господствующие высоты. Такие приспособленные организмы называются специализированными. Их судьба незавидна. Углубившись в «подгонку» себя к господствующей среде, холодной (как зверозубые пресмыкающиеся) или теплой (как динозавры), они сожгут за собой эволюционные мосты и с переменой вектора развития среды (с похолодания на потепление или наоборот) окажутся беспомощными и вымрут (как угасли зверозубые рептилии и динозавры).
«Отыграть эволюцию назад» им не позволит закон Долло о необратимости эволюции, а эволюция необратима потому, что не является прямолинейной, и угадать в ней извилистые пути отступления в полном соответствии с путями наступления – нереально. Например, мы помним, что с похолоданием в ранней юре двуногие прозауроподы встали на четвереньки и сделались могучими зауроподами. Но переделать свою прожорливую мезозойскую природу они не смогли и с необратимым похолоданием в конце мела сошли с эволюционной сцены. То есть они сумели отыграть эволюцию назад лишь отчасти и поплатились за это местом под солнцем.
Напротив, организмы, воздержавшиеся от углубленного приспособления к наличной среде, не пережившие подходящих количестве
Date: 2015-09-03; view: 460; Нарушение авторских прав