Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Углеводы
|
|
Углеводы – это полиатомные альдегидо- или кетоспирты, которые подразделяются в зависимости от количества мономеров на моно-, олиго- и полисахариды.
Таблица 3.3. Основные представители углеводов
| Углеводы | Основные представители |
| Моносахариды | Гексозы (глюкоза, манноза, галактоза, фруктоза), триозы, тетрозы, пентозы |
| Производные моносахаридов | Сахарные кислоты, дезоксисахара, аминосахара, сиаловые кислоты |
| Дисахариды | Мальтоза, лактоза, сахароза |
| Олигосахариды | Мальтодекстрин |
| Полисахариды: | |
| -гомополисахариды | Крахмал, гликоген, целлюлоза |
| -гетерополисахариды | Гликозаминогликаны |
Моносахариды (глюкоза, фруктоза, галактоза и др.), олигосахариды (сахароза, мальтоза, лактоза) и перевариваемые полисахариды (крахмал, гликоген) являются основными источниками энергии, а также выполняют пластическую функцию (табл. 3.3).
Неперевариваемые полисахариды, или пищевые волокна, играют в питании важнейшую роль, участвуя в формировании каловых масс, регулируя моторную функцию кишечника, выступая в качестве сорбентов (табл. 3.4). Разделяют нерастворимые полисахариды: целлюлоза, гемицеллюлоза; растворимые: каррагинаны, альгинаты, арабиногалактан и др. Именно растворимые полисахариды являются пробиотиками. К пищевым волокнам относят и не являющийся углеводом лигнин.
Таблица 3.4. Роль неперевариваемых полисахаридов (пищевых волокон) в питании
| Пищевые волокна | Основные эффекты |
| Прием пищи | увеличение объема пищи и периода ее приема снижение энергетической плотности пищи усиление чувства насыщения |
| Влияние на верхние отделы желудочно-кишечного тракта | торможение опорожнения желудка стимуляция процессов желчеотделения |
| Влияние на тонкую кишку | связывание нутриентов, торможение абсорбции глюкозы, аминокислот и холестерина, токсических веществ торможение гидролиза крахмала |
| Влияние на толстую кишку | нормализация состава кишечной микрофлоры формирование каловых масс и повышение скорости их транзита |
Перевариваемые углеводы в тонком кишечнике расщепляются до дисахаридов, а далее, путем пристеночного пищеварения, до моносахаридов (рис. 3.2).
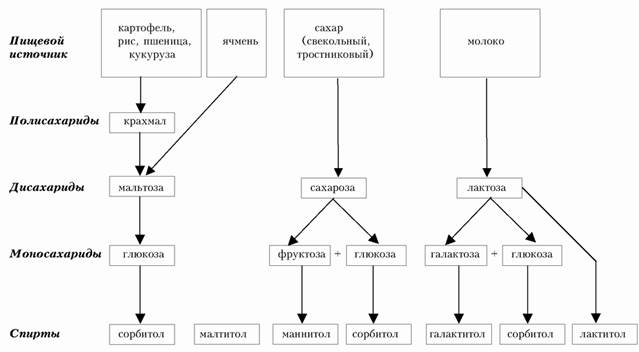
Рис. 3.2. Обмен пищевых растворимых углеводов (схема).
Всасывание моносахаров происходит путем облегченной дифузии и активного транспорта, что обеспечивает высокую их абсорбцию даже при низкой концентрации в кишечнике. Основным углеводным мономером является глюкоза, которая по системе воротной вене доставляется в печень, а далее или метаболизируется в ней, или поступает в общий кровоток и доставляется в органы и ткани.
Метаболизм глюкозы в тканях начинается с образования глюкозо-6-фосфата, который, в отличие от свободной глюкозы, не способен покидать клетку. Дальнейшие превращения этого соединения идут в следующих направлениях:
– расщепление вновь до глюкозы в печени, почках и эпителии кишечника, что позволяет поддерживать постоянный уровень сахара в крови;
– синтез депонируемой формы глюкозы – гликогена – в печени, мышцах и почках.
– окисление по основному (аэробному) пути катаболизма;
– окисление по пути гликолиза (анаэробного катаболизма), обеспечивающего энергией интенсивно работающие (мышечная ткань) или лишенные митохондрий (эритроциты) ткани и клетки;
– по пентозофосфатному пути превращений, происходящему под действием коферментной формы витамина B1, в ходе которого генерируются продукты, используемые в синтезе биологически значимых молекул (НАДФ•Н2, нуклеиновых кислот).
Таким образом, метаболизм глюкозы может происходить по различным направлениям, использующим ее энергетический потенциал, пластические возможности или способность депонироваться.
Обеспечение тканей глюкозой, как энергетическим материалом, происходит за счет экзогенных сахаров, использования запасов гликогена и синтеза глюкозы из неуглеводных предшественников. В базальном (доабсорбционном) состоянии печень вырабатывает глюкозу со скоростью, равной ее утилизации во всем организме. Примерно 30% производства глюкозы печенью происходит за счет гликогенолиза, а 70% – как результат глюконеогенеза. Общее содержание гликогена в организме составляет примерно 500 г. Если нет экзогенного поступления глюкозы, его запасы истощаются через 12–18 часов. При отсутствии резервного гликогена в результате голодания резко усиливаются процессы окисления другого энергетического субстрата – жирных кислот. Одновременно увеличивается скорость глюконеогенеза, направленного в первую очередь на обеспечение глюкозой головного мозга, для которого она является основным источником энергии.
Синтез глюкозы происходит из аминокислот, лактата, пирувата, глицерина и жирных кислот с нечетной углеродной цепью. Большинство аминокислот способны быть предшественниками глюкозы, однако основную роль при этом, как сказано выше, играет аланин. Из аминокислотных источников происходит синтез примерно 6% эндогенной глюкозы, из глицерина, пирувата и лактата соответственно 2%, 1% и 16%. Вклад жирных кислот в глюконеогенез малозначим, поскольку лишь небольшой процент их имеет нечетное углеродное число.
В постабсорбционном состоянии печень из органа, вырабатывающего глюкозу, превращается в запасающий. При повышении концентрации глюкозы скорость ее утилизации периферическими тканями почти не изменяется, поэтому основным механизмом элиминации ее из кровотока является депонирование. Только небольшая часть избыточной глюкозы непосредственно участвует в липогенезе, который происходит в печени и в жировой ткани. Эти особенности углеводного метаболизма становятся значимыми при парентеральном введении высококонцентрированных растворов глюкозы.
Обмен глюкозы в мышцах по сравнению с печенью носит редуцированный характер. Ведь печень обеспечивает углеводами все органы и ткани, а мышцы работают в соответствии с принципом самообслуживания. Здесь происходит создание запаса гликогена в состоянии покоя и использование его и вновь поступающей глюкозы при работе. Запасы гликогена в мышцах не превышают 1% от их массы. Основные энергетические потребности интенсивно работающей мускулатуры удовлетворяются за счет окисления продуктов обмена жиров, а глюкоза используется здесь в гораздо меньшей степени. В процессе гликолиза из нее образуется пируват, который утилизируют скелетные мышцы. При повышении уровня работы мышечная ткань вступает в анаэробные условия, производя пируват в лактат. Тот диффундирует в печень, где используется для глюкозного ресинтеза, а также может окисляться в миокарде, который практически всегда работает в аэробных условиях.
Регуляция содержания глюкозы в крови осуществляется гормонами: инсулином, глюкагоном, глюкокортикостероидами, адреналином, соматостатином. Инсулин играет ключевую роль в регуляции углеводного метаболизма, обеспечивая поступление глюкозы в клетку, активируя ее транспорт через клеточные мембраны, ускоряя окисление. Кроме того, он стимулирует гликогенообразование, липо- и протеиногенез. Одновременно тормозится гликогенолиз, липолиз и глюконеогенез. Глюкагон, наоборот, активирует процессы, ведущие к росту концентрации глюкозы в крови. Глюкокортикостероиды действуют в направлении гипергликемии, стимулируя процессы продукции глюкозы печенью. Адреналин усиливает мобилизацию гликогена. Соматотропный гормон увеличивает секрецию и глюкагона, и инсулина, что ведет как к увеличению депонирования глюкозы, так и к усилению утилизации. Соматостатин тормозит продукцию соматотропина и опосредованно сдерживает выработку инсулина и глюкагона.
Специфические превращения других перевариваемых углеводов по сравнению с глюкозой имеют меньшее значение, поскольку в основном их метаболизм происходит через образование глюкозы. Отдельное значение придается фруктозе, которая также является быстроутилизируемым источником энергии и еще легче, чем глюкоза, участвует в липогенезе. При этом утилизация не перешедшей в глюкозо-фосфат фруктозы не требует стимуляции инсулином, соответственно она легче переносится при нарушении толерантности к глюкозе.
Пластическая функция углеводов заключается в их участии в синтезе гликопротеинов и гликолипидов, а также возможности выступать предшественниками триглицеридов, заменимых аминокислот, использоваться при построении многих других биологически значимых соединений.
ЖИРЫ
Жиры (липиды), представленные в организме в основном триглицеридами (соединениями глицерина и жирных кислот), представляют собой наиболее важный энергетический субстрат. Благодаря высокой калорической "плотности" (в среднем 9 ккал/г, по сравнению с 4 ккал/г у глюкозы), жиры составляют более 80% энергетических запасов в организме. Энергетическая ценность отдельных триглицеридов определяется длиной углеродных цепей жирных кислот, поэтому при использовании специализированных энтеральных и парентеральных продуктов их калорийность может быть ниже средней (например, у препаратов триглицеридов со средней углеродной цепью – 8 ккал/г). При нормальном питании жиры обеспечивают до 40% от общей калорийности питания.
Жирные кислоты подразделяются на насыщенные и ненасыщенные (содержащие двойные химические связи). Источником насыщенных жирных кислот является преимущественно животная пища, ненасыщенных продукты растительного происхождения.
Пищевая ценность жировых продуктов определяется их триглицеридным спектром и наличием других факторов липидной природы. Синтез насыщенных и мононенасыщенных жирных кислот возможен в организме человека.
Особое значение в диетологии придается ненасыщенным жирным кислотам, являющимся эссенциальными факторами питания. Полиненасыщенные жирные кислоты, несущие в организме важнейшие функции (это предшественники ряда биологически активных веществ), должны поступать экзогенно. К эссенциальным жирным кислотам относятся линолевая и линоленовая. Линолевая кислота метаболизируется в организме в арахидоновую, а линоленовая в эйкозапентайеновую кислоты, которые могут поступать в организм с мясными и рыбными продуктами, но в незначительных количествах (табл. 3.5). компонентами клеточных мембран, предшественниками гормоноподобных веществ. Линолевая и образуемая из нее арахидоновая кислота относятся к ω- 6 -жирным кислотам, линоленовая кислота и продукты ее метаболизма эйкозопентаеновая и дезоксогексаеновая – ω- 3 -жирные кислоты. Дефицит эссенциальных жирных кислот в рационе вызывает, прежде всего, нарушение биосинтеза арахидоновой кислоты, которая входит в большом количестве в состав структурных фосфолипидов и простагландинов. Содержание линолевой и линоленовой кислот во многом определяет биологическую ценность пищевых продуктов. Недостаточность эссенциальных жирных кислот развивается в основном у больных, находящихся на полном парентеральном питании без применения жировых эмульсий.
Таблица 3.5. Основные пищевые источники различных жирных кислот
| Жирные кислоты | Основные пищевые источники |
| Полиненасыщенные жирные кислоты | |
| ω- 6 | |
| линолевая | Овощное масло, орехи, семечки |
| ω- 3 | |
| линоленовая | Льняное, соевое масло |
| эйкозопентаеновая, дезоксогексаеновая | Рыба, рыбий жир, планктон |
| Мононенасыщенные жирные кислоты, ω- 9 | |
| олеиновая (цис-форма) | Растительные масла (оливковое, соевое, подсолнечное), орехи, авокадо |
| олеиновая (транс-форма) | Маргарины |
| Насыщенные жирные кислоты | Продукты животного происхождения |
При обработке растительных масел – создании маргаринов – происходит изомеризация ненасыщенных жирных кислот с созданием транс-изомеров, которые утрачивают некоторые биологические функции своих предшественников.
Триглицериды со средней длиной углеродной цепи (МСТ, СЦТ) имеют более высокую усвояемость, чем другие виды триглицеридов. Они гидролизируются в кишечнике без участия желчи, больше атакуются липазами. Кроме того, введение среднецепочечных триглицеридов оказывает гипохолестеринемический эффект, так как они не участвуют в мицеллообразовании, необходимом для всасывания холестерина. Недостатком применения препаратов, содержащих триглицериды со средней длиной углеродной цепи, является то, что они используются исключительно как энергетический (но не пластический) субстрат. Кроме того, окисление таких жирных кислот приводит к интенсивному накоплению кетоновых тел и может усугубить ацидоз.
Стерины и фосфолипиды не относятся к эссенциальным факторам питания, но играют важнейшую роль в метаболизме.
Фосфолипиды являются незаменимыми компонентами организма. Основная их роль – обеспечение фундаментальной структуры мембраны, как барьера проницаемости. Биосинтез структурных фосфолипидов в печени направлен на обеспечение ими самой печени и других органов. Фосфолипиды оказывают липотропное действие, способствуя мицеллообразованию жиров в пищеварительном тракте, транспорту их из печени, а также и стабилизируя липопротеины.
Стерины в животных продуктах представлены холестерином, а в растительных – смесью фитостеринов.
Холестерин является структурным компонентом мембран и предшественником стероидов (гормонов, витамина D, желчных кислот). Пополнение запасов холестерина происходит за счет кишечной абсорбции и биосинтеза (1 г в сутки). Количество всасывающегося в кишечнике холестерина ограничено (0,3–0,5 г в сутки), и при излишнем содержании в пище он выводится с фекалиями. Абсорбция холестерин ингибируется его растительными структурными аналогами фитостеринами. Сами фитостерины тоже могут включаться в эндогенные липидные образования, но их участие минимально. При избыточном поступлении холестерина с пищей его синтез в печени, кишечнике и коже практически прекращается.
Поступающий из кишечника в составе хиломикронов холестерин в значительной степени задерживается в печени, где используется для построения мембран гепатоцитов и в синтезе желчных кислот. В составе желчи в результате реабсорбции в организм возвращается около 40% жиров. Не подвергшиеся обратному всасыванию в кишечнике холестерин и желчные кислоты – это основной путь выведения холестерина из организма. В кровотоке липиды существуют в составе транспортных форм: хиломикронов, липопротеидов очень низкой плотности (ЛПОНП), липопротеидов низкой плотности (ЛПНП) и липопротеидов высокой плотности (ЛПВП). В энтероцитах образуются хиломикроны и ЛОПНП, в гепатоцитах – ЛПОНП и ЛПВП, в плазме крови – ЛПВП и ЛПНП (рис. 3.3).
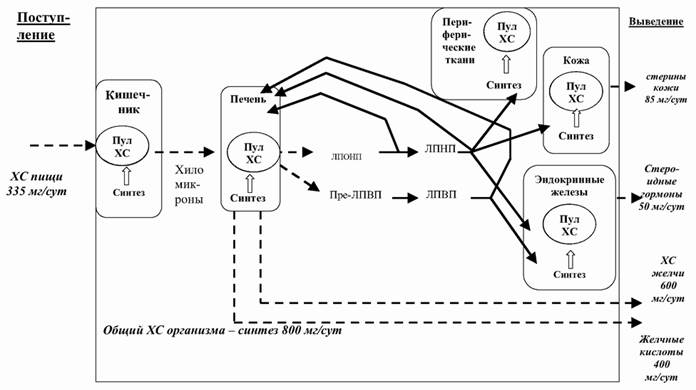
Рис. 3.3. Обмен холестерина (ХС) (по: I. M. Arias et al., 1982) (схема).
Хиломикроны и ЛПОНП транспортируют преимущественно триглицериды, а ЛПНП и ЛПВП – холестерин. Холестерин-содержащие липопротеиды регулируют баланс холестерина в клетках: ЛПНП обеспечивают потребности, а ЛПВП предупреждают избыточное накопление.
Различают пять типов дислипопротеинемий. I тип связан с нарушением лизиса хиломикронов, IIа тип – результат нарушения распада ЛПНП и снижения поступления холестерина в клетку, II тип характеризуется замедлением распада ЛПОНП, IV тип связан с усилением синтеза триглицеридов в печени в результате гиперинсулинизма, механизмы развития IIб и V типов точно не известны.
На состав триглицеридов и липопротеинов выраженное влияние оказывает состав пищи. Продукты животного происхождения, включающие преимущественно полиненасыщенные жирные кислоты и холестерин, имеют атерогенный эффект, содержание в крови ЛПВП и триглицеридов. Наоборот, ненасыщенные жирные кислоты (их источник растительные масла) и в особенности ω-3-жирные кислоты (содержащиеся в жире рыб) оказывают профилактическое действие (табл. 3.6).
Таблица 3.6. Влияние жирных кислот на липопротеидный спектр
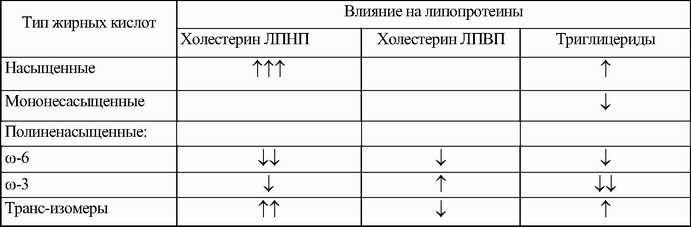
Примечание:↑ – повышают, ↓ – снижают.
Как и при метаболизме углеводов, ведущую роль в липидном обмене играет печень. Исключительно в печени локализованы такие процессы, как биосинтез холестерина, желчных кислот и фосфолипидов. В обмене других липидов ей присущи модифицирующие и регулярные функции. В отличие от богатых запасов гликогена, печень практически не содержит собственных резервов триглицеридов (менее 1%), однако занимает ключевую позицию в процессах мобилизации, потребления и синтеза жиров в других тканях.
Такая ее роль основана на том, что практически все потоки обмена жиров проходят через печень: липиды пищи в виде хиломикронов поступают в нее через общий кровоток по печеночной артерии; свободные жирные кислоты, мобилизованные из жировых депо переносятся в виде комплексов с альбумином; соли желчных кислот, реабсорбируясь в кишечнике, вновь поступают по воротной вене.
Энергетический потенциал липидов обеспечивает более половины основной энергетической потребности большинства тканей, что особенно выражено в условиях голода. При голодании или снижении утилизации глюкозы, триглицериды жировой ткани гидролизируются в жирные кислоты, которые в таких органах как сердце, мышцы и печень подвергаются интенсивному β-окислению с образованием АТФ.
Продуктами неполной утилизации жиров печенью являются кетоновые тела. К ним относятся ацетоуксусная кислота, β-оксибутират и ацетон. В норме кетоны образуются в незначительном количестве и полностью утилизируются как источник энергии нервной тканью, скелетными и висцеральными мышцами. В условиях ускоренного катаболизма жирных кислот и/или снижения утилизации углеводов синтез кетонов может превысить возможности их окисления внепеченочными органами и привести к развитию метаболического ацидоза. Ингибирующее влияние на кетоногенез оказывают углеводы рациона.
Головной мозг и нервная ткань практически не используют жиры как источник энергии, так как здесь не происходит β-окисления. Однако эти ткани могут использовать кетоновые тел. В норме доля процессов окисления кетоновых тел незначительна по сравнению с катаболизмом глюкозы. Однако в условиях голодания кетоновые тела становятся важным альтернативным источником энергии.
Кетоны используются и мышцами, наряду с происходящей здесь утилизацией глюкозы и β-окислением. При незначительной физической нагрузке мышцы окисляют в основном углеводы, увеличение интенсивности и длительности работы требует преобладания катаболизма жиров, β-окисление в большинстве тканей стимулируется переносчиком липидов карнитином, но особенно весомое значение он имеет для мышечной ткани.
Свободнорадикальные формы кислорода вызывают процессы перекисного окисления, которому в первую очередь подвержены полиненасыщенные жирные кислоты. Это физиологический процесс, обеспечивающий регуляцию активности клеток. Однако при избыточном образовании свободных радикалов их окислительная активность приводит к нарушению структуры и гибели клетки. Для ограничения перекисного окисления существует система антиоксидантной защиты, которая ингибирует образование свободных радикалов и разлагает токсичные продукты их окисления. Функционирование этой системы во многом зависит от алиментарно поступающих антиоксидантов: токоферолов, селена, серосодержащих аминокислот, аскорбиновой кислоты, рутина.
Синтез жирных кислот (за исключением эссенциальных) может происходить из любых веществ, для которых конечным продуктом метаболизма является ацетил-Ко-А, но основным источником липогенеза являются углеводы. При излишнем количестве глюкозы в печени (после еды) и достаточных запасах гликогена глюкоза начинает разлагаться до предшественников жирных кислот. Если потребление углеводов превышает энергетические потребности организма, то их избыток в дальнейшем превращается в жиры.
Регуляция метаболизма жирных кислот и глюкозы тесно связаны: повышенное окисление жирных кислот ингибирует утилизацию глюкозы. Поэтому инфузия жировых эмульсий с соответственным повышением уровня свободных жирных кислот в крови ослабляет действие инсулина на утилизацию глюкозы и стимулирует печеночный глюконеогенез. Этот момент немаловажен при парентеральном питании больных с изначально нарушенной толерантностью к глюкозе.
Взаимосвязь между обменом основных нутриентов осуществляется за счет существования общих предшественников и промежуточных продуктов метаболизма. Наиболее важным общим продуктом метаболизма, участвующим во всех обменных процессах, является ацетил-Ко-А. (рис. 3.4) Поток веществ в сторону липогенеза от углеводных и белковых источников через ацетил-Ко-А носит однонаправленный характер, поскольку в организме не существует механизма, обеспечивающего превращение этого двухуглеродного вещества в трехуглеродные соединения, необходимые для глюконеогенеза или синтеза заменимых аминокислот. Хотя при катаболизме липидов и происходит образование небольших количеств промежуточных трехуглеродных продуктов, оно малозначительно.
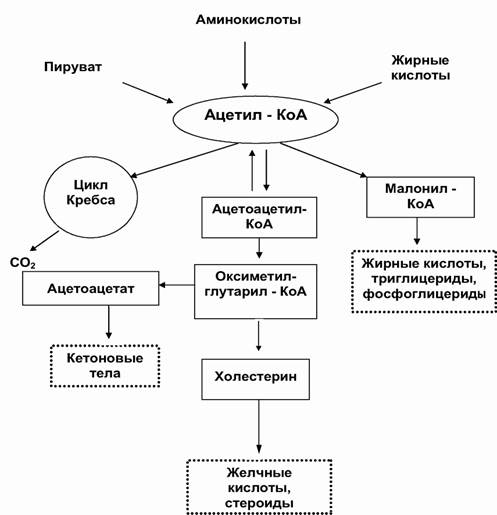
Рис. 3.4. Основные взаимоотношения в метаболизме липидов (по Бышевскому А. Ш., Терсенову О.А. 1994; схема).
Общим конечным путем всех метаболических систем является цикл Кребса и реакции дыхательной цепи. Цикл лимонной кислоты является поставщиком двуокиси углерода для реакций синтеза жирных кислот и глюконеогенеза, образования мочевины и пуринов и пиримидинов. Взаимосвязь между процессами углеводного и азотного обмена достигается посредством промежуточных продуктов цикла Кребса. Другие звенья этого цикла являются предшественниками липонеогенеза.
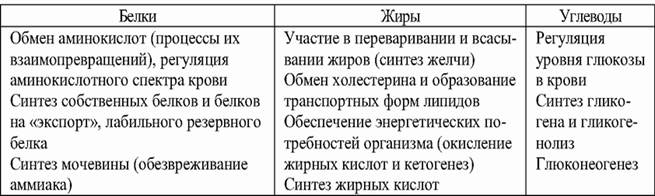
Основную роль в метаболизме нутриентов играет печень (табл. 3.7).
Таблица 3.7. Роль печени в метаболизме белков, жиров и углеводов
Date: 2015-07-02; view: 1771; Нарушение авторских прав