Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Электрическая активность коры
|
|
Деятельность больших полушарий головного мозга сопровождается генерацией ее электрической активности. Наряду с вызванной электрической активностью в коре существует фоновая (основная) электрическая активность, колебания которой происходят постоянно, без специального воздействия. Фоновая электрическая активность включает широкий спектр электрических колебаний разной продолжительности, от квазипостоянного потенциала коры и сверхмедленных колебаний потенциалов до быстрых потенциалов действия. Между поверхностью коры и белым веществом имеется устойчивая квазипостоянная разница потенциалов: поверхность коры несет более положительный заряд, чем глубокие слои. Этот постоянный потенциал возникает вследствие разной степени поляризации дендритного и аксонного полюсов пирамидных нейронов коры, ось которых вертикально ориентирована. При естественном сне, наркозе и других изменениях состояния потенциал смещается в положительную сторону; при пробуждении, активации — в отрицательную. От коры регистрируются также сверхмедленные колебания потенциалов с секундными, декасекундными, минутными и много-
|

|
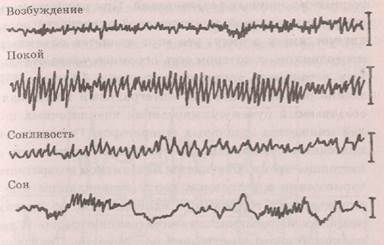
минутными периодами. Сверхмедленные колебания потенциалов отражают деятельность медленной регу- ляторной системы мозга; их природа связана с различием ионного состава по обе стороны гематоэнцефа- лического барьера, поляризацией нейроглии, колебаниями ионного состава межклеточного пространства. Наиболее изученными и широко регистрируемыми для оценки функционального состояния являются медленные колебания фоновой электрической активности, которые можно регистрировать непосредственно от колеи головы животных (Правдич-Неминский, 1929) и человека (Бергер, 1929). Запись медленных электрических колебаний от кожи головы — электроэнцефалограмма (ЭЭГ) — содержит все те же виды колебаний, что и запись непосредственно от коры — элект-рокортикограмма (ЭКоГ). Характер медленной электрической активности очень изменчив и тонко реагирует на сдвиги функционального состояния и уровень мозговой деятельности (рис. 8.4). В состоянии спокойного бодрствования в ЭЭГ хорошо выражены альфа-волны с частотой 8-12 колебаний в секунду. При закрывании глаз их представленность возрастает, они переходят в регулярный альфа-ритм, более выраженный в затылочной и теменных областях. В состоянии активного бодрствования, деятельности или напряжения альфа-волны сменяются бета-волнами с частотой 18-35 колебаний в секунду, лучше выраженными в лобных и височных областях. При снижении уровня бодрствования и наступлении сна выражены тега-волны с частотой от 4 до 7 колебаний в секунду, а при глубоком сне или поражениях тканей мозга — дельта-волны с частотой 0,5-3,5 колебаний в секунду. Выделяют также волны гамма-ритма с частотой свыше 35 и сигма-ритма с частотой 13—18 колебаний в секунду и веретенообразно изменяющейся амплиту-
Рис. 8.4. Изменения ЭЭГ при смене функциональных состояний головного мозга (Оке, 1969)
дои. Гамма-волны усиливаются при двигательной активности и напряжении, сигма-волны — в дремотном состоянии и при действии барбитуратов. При изменении функционального состояния головного мозга изменяется диапазон доминирующих ритмов ЭЭГ. Происхождение волн ЭЭГ связано с медленной электрической активностью преимущественно поверхностных структур больших полушарий. В происхождение медленной электрической активности вносят вклад в первую очередь постсинаптические потенциалы сомы и дендритов пирамидных нейронов коры, а также потенциалы нейроглии, сдвиги ионного состава межклеточных пространств, деполяризация сети
тонких дендритных разветвлений. При рассмотрении природы медленной электрической активности коры следует иметь в виду, что мозг является объемным проводником, в котором есть огромное число отдельных источников электрического тока. Поэтому ЭЭГ можно рассматривать как интегральный потенциал, создаваемый путем суммирования внеклеточных полей множества отдельных генераторов. Ритмичность колебаний суммарной электрической активности в настоящее время объясняется механизмом возвратного торможения в нейронных цепях, приводящим к периодическому торможению одновременно во многих нейронах и ритмическому чередованию гипер- и де-поляризационных колебаний потенциалов. Показано, что происхождение альфа-ритма и сигма-ритма обусловлено системами возвратного торможения в специфических и неспецифических структурах таламуса. Существует большое число различных методов анализа ЭЭГ, позволяющих оценивать разнообразные изменения состояния больших полушарий головного мозга при разных условиях, бодрствовании и сне, внимании и релаксации, активации и торможении, в норме и при патологии. Наиболее полно изменения функционального состояния отражаются в параметрах пространственно-временной организации фоновой электрической активности больших полушарий головного мозга. Вызванная электрическая активность представляет собой изменения фоновой электрической активности или кратковременные, вызванные потенциалы в ответ на воздействие. Изменение фоновой ЭЭГ у человека происходит при использовании функциональных проб: 1) открыванием и закрыванием глаз; 2) ритмическими световыми мельканиями или звуковыми толчками с разной частотой (3-20 в с); 3) выполнением арифметических и логических заданий; 4) гипервен-
тиляцией, т.е. усилением дыхания в течение 3-5 мин (рис. 8.5). Наиболее частный характер изменений ЭЭГ при открывании глаз и других пробах состоит в «де-синхронизации» электрической активности, когда хорошо выраженные медленные колебания альфа-

Рис. 8.5. Вызванные потенциалы коры головного мозга кошки при электростимуляции контралатерального седалищного нерва (по Куллаяде, 1968). Первичные ответы: 1,2 — корот-колагпентные; 9, 10 — длиннолатентные. Ранние отрицательные реакции: 3, 4, — постсинаптические; 5,6 — преси-наптические I типа; 7,8 — пресинаптические II типа. Отклонение луча вниз соответствует положительному колебанию потенциала. В центре — схема мозга кошки с системой стереотаксических координат и указанием точек отведения потенциалов
ритма сменяются низкоамплитудными высокочастотными колебаниями бета-ритма. Вызванные потенциалы, регистрируемые с поверхности коры, разделяют на первичные и вторичные ответы. Первичный ответ — это кратковременная (20-30 мс) электрическая реакция коры на залп импульсов из специфических афферентов, возникающая в определенных участках их проекции. Первичный ответ, регистрируемый с поверхности коры, представляет позитивно-негативное колебание, которому предшествуют небольшие быстрые колебания пресинаптического происхождения. Позитивная фаза первичного ответа отражает местное возбуждение сомы пирамидных нейронов III и IV слоев коры, а негативная фаза — синаптическое возбуждение апикальных дендритов в I слое коры по афферентам пирамидных клеток или прямо от неспецифических афферентов. Функциональное значение первичных ответов может быть связано с выделением начального момента действия раздражителя. Вторичные ответы могут быть локальными или иметь широкую предетавленностъ, возникать с разным латентным периодом, обычно большим, чем у первичных ответов. К локальным вторичным ответам с коротким латентным периодом относятся ранние отрицательные реакции, ранние ассоциативные и каллозаль-ные ответы. Вторичные генерализованные ответы с большим латентным периодом ретикулокоркового и таламокоркового происхождения более чувствительны к изменениям функционального состояния мозга. Вызванные потенциалы, регистрируемые с поверхности кожи головы человека, имеют несколько негативных и позитивных колебаний с разной пиковой ла-тентностыо (от 50 до 300 мс). Они являются отражением процессов не прихода информации в кору, а ее обработки, и значительно изменяются в связи со сдви-
гами уровня внимания, степени умственного напряжения, эмоциональной оценки стимула.
Date: 2016-11-17; view: 535; Нарушение авторских прав