Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Глава 1 роль полового размножения в эволюции
|
|
Способы размножения
Размножение — важнейший биологический процесс, обеспечивающий поддержание и увеличение численности вида, возможность его расселения и, в конечном итоге, успех борьбы за существование. В животном мире существует целый ряд способов размножения, например, прямое деление, характерное для низших беспозвоночных, а также партеногенез, отмечаемый даже у высших позвоночных. Совершенно очевидно, что именно бесполое размножение представляет собой наиболее простой и наименее энергоемкий путь увеличения численности особей. Однако, почему—то, в процессе эволюции возник сложный, сопряженный с множеством проблем и условностей, процесс полового размножения.
Половое размножение появляется у животных уже на самых низших ступенях эволюционной лестницы. Так, уже у простейших одноклеточных микроорганизмов — инфузорий, размножающихся прямым делением, наблюдается так называемая конъюгация, представляющая собой некий аналог полового процесса. У целого ряда беспозвоночных, а также многих позвоночных животных отмечается такое явление как партеногенез, при котором самки откладывают яйца или рожают живых детенышей без участия самцов. Вылупляются из этих яиц или рождаются, подобным образом только самки, причем, интересно, что в природе существуют целые локальные популяции подобных видов. В экспериментальных условиях партеногенез возможен даже у млекопитающих.
У целого ряда беспозвоночных животных имеет место гермофрадитизм, при котором у каждой особи имеются как мужские так и женские половые железы. Среди наиболее известных животных гермафродитами являются, например, дождевые черви, пиявки и многие виды улиток. У подобных животных каждая особь производит и яйцеклетки и сперматозоиды, они спариваются друг с другом, производя взаимный обмен половыми клетками. При этом, несмотря на наличие у каждой особи половых клеток обоих типов, они крайне редко прибегают к самооплодотворению, и напротив, обычно стремятся к спариванию с другими, предпочтительно не родственными особями.
У таких беспозвоночных, как кишечнополостные, черви разных типов, иглокожие, параллельно существуют как половое так и бесполое размножение. Для множества беспозвоночных характерно чередование половых и бесполых поколений. У некоторых видов наблюдается чередования партеногенеза с нормальным половым размножением. Так, например, тли, в течение всего лета размножаются живорождением, при котором, неоплодотворенные самки рожают одних самок. Молодые самки тлей начинают рожать следующих самок уже в возрасте нескольких часов. Таким образом, в течение лета проходит огромная череда бесполых поколений. Зоологи подсчитали, что если бы все потомство одной тли выживало, то оно за одно лето могло бы покрыть сплошным покрывалом весь Земной шар. К счастью, поскольку тлями питается множество других животных, этого не происходит. Казалось бы, данный способ размножения тлей весьма успешен и не требует желать ничего лучшего. Однако, осенью многие тли откладывают яйца из которых выводятся самцы, которые оплодотворяют сравнительно небольшое количество самок. В то время как осенью большинство тлей, закончив свой жизненный цикл, погибает, оплодотворенные самки успешно переживают зиму, и весной откладывают яйца. Из яиц вылупляются крылатые тли, они разлетаются в разных направлениях, и, осев на подходящих кормовых растениях, утрачивают крылья и начинают размножаться партеногенезом. Очевидно, что для успешного выживания в условиях континентального климата тлям, оказывается необходимым расширение спектра изменчивости в геноме, обеспечиваемое половым размножением.
Таким образом, основное значение полового размножения заключается не просто в увеличении количества особей, а в расширении генофонда, в дальнейшем способствующему естественному отбору.
При половом размножении в популяции создается более высокая генетическая изменчивость. В результате целого ряда процессов, гены, носителями которых изначально были родители, оказываются в новой комбинации в потомках. Именно благодаря рекомбинации внутри помета обнаруживаются многочисленные генетические различия, что повышает адаптационный потенциал популяции и вида в целом.
Зачем нужны два пола?
В процессе эволюции мужские и женские организмы играют разные роли. Проблему неравноценности полов подробно исследовал российский ученый В.А. Геодакян, который показал целесообразность этого явления для существования вида. На всех стадиях полового процесса мужской пол подвергается более жесткому воздействию естественного отбора. Известно, что на всех этапах онтогенеза смертность самцов выше, чем самок. Этот процесс начинается фактически с момента оплодотворения, в процессе которого участвуют миллионы сперматозоидов и считанные единицы яйцеклеток. Показано, что среди эмбрионов ранних стадий развития значительно преобладают мужские. Даже, несмотря на то, что в период эмбрионального развития мужских эмбрионов гибнет больше, чем женских, доля рожденных самцов млекопитающих несколько превышает долю рожденных самок. Анализ пороков развития эмбрионов и новорожденных показал, что для самок характерны пороки развития атавистического характера, в то время как для самцов характерны оригинальные пороки, носящие футуристический характер и являющиеся, как бы, пробами и ошибками эволюции. К моменту прекращения молочного вскармливания количество самцов и самок в пометах обычно уравновешивается. В следующие возрастные периоды самцы гибнут как от разнообразных пороков развития, так и вследствие более высокой, чем у самок активности.
Таким образом, женский пол обеспечивает количественную сторону процесса — чем больше самок, тем выше для вида шансы для выживания и размножения. Женский пол сохраняет консервативные программы вида и обеспечивает их передачу потомству. Поэтому биологически выгодно, чтобы женские особи были менее изменчивы и более устойчивы ко всяким воздействиям. Этому же способствует и то, что у подавляющего большинства видов самки не имеют яркой окраски и других украшений, характерных для самцов, что делает их менее заметными для конкурентов и хищников. Мужским особям свойственна более высокая изменчивость, которая обеспечивает выживаемость вида и его адаптацию к условиям среды обитания. Самцы первыми реагируют на неблагоприятные воздействия, часть из них погибает. Оставшиеся в живых передают свои признаки, оказавшиеся полезными в данных условиях, потомству. Потеря самок сказывается главным образом на возможном числе потомков. Элиминирование наименее приспособленных к данным условиям особей способствует изменениям качества потомства в нужном направлении, и приносит пользу виду, способствуя его адаптации, обеспечивая эволюцию.
Таким образом, самцы представляют собой как бы модель, на которой природа испытывает различные изменения, могущие иметь значение для эволюции вида. Они представляют лабильную часть популяции, ее передовой отряд, продвинутый далеко вперед навстречу факторам среды. Между ними и золотым ядром популяции — самками существует определенная дистанция, необходимая для отбора из всего разнообразия самцов наиболее удачных и достойных для включения в генофонд вида (Акоев, Алексеева, 1985). Именно поэтому в размножении принимает участие лишь малая толика самцов от числа родившихся. Основная часть самцов в природе обречена на безбрачие.
В популяциях стадных животных кроме основных, активно участвующих в размножении, всегда существует некий резерв самцов, которые обычно держаться на периферии или несколько поодаль от основного стада. Так на лежбищах морских котиков и моржей, кроме самцов владеющих «гаремами» самок всегда присутствует довольно много холостых самцов, называемых периферийными. Когда владельцы гаремов, истощенные голоданием, драками и спариванием, через 20–30 дней бросают самок и уходят в холостяцкие стада, их места тотчас занимают перифирийные самцы.
Для насекомых, например, типично появление ограниченного контингента самцов лишь в те жизненные моменты, когда необходимо оплодотворение самок. Весь их жизненный цикл сводится именно к данному процессу, после чего они погибают. Подобное явление наблюдается, в частности, у общественных насекомых. Гибель самцов после периода размножения происходит часто и у многих полигамных видов млекопитающих. Так, например, погибают или полностью утрачивают функции доминантов самцы сайгаков, ослабевшие после гона, вследствие усиленной половой нагрузки, вынужденной голодовки и драк с соперниками. Самцы одного из мелких видов сумчатого животного — бурой сумчатой мыши неминуемо гибнут с симптомами тяжелого стресса после очень краткого периода гона, длящегося 1,5–2 суток, в течение которого один самец оплодотворяет до двух десятков самок. Самцов многих беспозвоночных, например, пауков и богомолов, сразу же после спаривания, а иногда даже во время него, поедают более крупные самки.
Различные роли самцов и самок в процессе размножения привели к возникновению в процессе эволюции различий, как во внутреннем, так и внешнем строении.
Половой отбор
Половой отбор представляет собой важную составляющую естественного отбора. Ч. Дарвин определял его как «форму естественного отбора, направленную на закрепление преимуществ особей, проявленных в период размножения». Ч. Дарвин сформулировал два основных принципа полового отбора, которые могут действовать в любой комбинации. С одной стороны, предполагалось, что существует непосредственная конкуренция между самцами в противоборствах и групповом доминировании. С другой стороны, он допускал и опосредованное соперничество через демонстрации индивидуальных особенностей и способностей к охране территории, строительству гнезд и заботе о потомстве.
Исходя из двух этих принципов, Ч. Дарвин предложил гипотезу происхождения полового диморфизма. По его мнению, половой диморфизм является следствием полового отбора, а первопричиной половых различий является конкуренция самцов за внимание самок. Самцы приобретают некоторые свойства, которые привлекают самок. Эти свойства сохраняются благодаря выбору самок, тогда как самки практически не подвергаются давлению полового отбора. В результате самцы приобретают яркую окраску, громкое пение и другие привлекающие внимание качества, тогда как самки — орудие половой эволюции — остаются невзрачными партнерами. При этом соотношение полов в популяции обычно приблизительно равно. Оно не всегда точно соблюдается, но общая тенденция не вызывает сомнений. Половой отбор, очевидно, был причиной происхождения сперматозоидов и яйцеклеток, а также различных стратегий размножения и заботы о потомстве. Конкуренция между самцами носит название внутриполового отбора. Соперничество за внимание самок принято называть межполовым отбором. Говоря о половом отборе, Ч. Дарвин отмечал, что основная его доля приходится на внутриполовой (внутрисамцовый) отбор. Самкам он отводил роль относительно пассивных выборщиц, которые отдают предпочтение самцам с наиболее привлекательными формами ухаживания, особенностями окраски или размерами. Однако, несмотря на кажущуюся пассивность, в ходе естественного отбора, ведущую роль играет именно выбор, осуществляемый самками.
Половой отбор начинается уже на уровне половых клеток. Так многие виды животных, обитающие в океанских глубинах, синхронно выбрасывают огромные количества яйцеклеток и сперматозоидов непосредственно в воду. Это обычно происходит в определенные фазы луны, когда океанский прилив достигает максимума. Шансы на оплодотворение при этом крайне малы, например, у коралловых полипов оплодотворяется примерно одна яйцеклетка на десять миллионов. У животных с внутренним оплодотворением существует жесткая конкуренция между сперматозоидами, количество которых на много порядков превышает количество яйцеклеток.
Жестокая конкуренция за право оплодотворения существует между самцами. При этом отбор идет в двух направлениях. С одной стороны, самцы ожесточенно сражаются друг с другом, отстаивая право сильнейшего, а с другой стороны, самки выбирают наиболее привлекательных, здоровых и осуществляющих самую правильную стратегию ухаживания. В брачный период многие животные образуют массовые скопления, которые способствуют ожесточению полового отбора.
ГЛАВА 2 БИОЛОГИЯ ПОЛА. ОПРЕДЕЛЕНИЕ ПОЛА
Пол — это совокупность признаков и свойств организма, обеспечивающих его участие в воспроизводстве потомства и передаче наследственной информации за счет образования половых клеток — гамет.
Генетические механизмы определения пола у собак
В кариотипе каждого животного существует пара хромосом, от которых зависит определение пола животного. У самок млекопитающих эти хромосомы одинаковы и представлены двумя половыми Х хромосомами. У самцов половые хромосомы различны, одна — Х, другая — Y. При этом набор аутосом или соматических хромосом у самцов и самок каждого вида одинаков. Таким образом кариотип собаки представлен 38 парами соматических и одной парой половых хромосом (XX у суки и ХY у кобеля). При образовании гамет в каждой из яйцеклеток оказывается по одной Х-хромосоме. Сперматозоиды же различны, половина из них несет Х-хромосому, а половина Y-хромосому.
Пол, имеющий две одинаковые половые хромосомы и продуцирующий гаметы одного типа (содержащие только Х-хромосомы), называется гомогаметным. Как уже упоминалось, у млекопитающих это самки, в то время как у птиц и насекомых — самцы.
Пол, имеющий две разные половые хромосомы и продуцирующий гаметы двух типов (с X- и Y-хромосомами), называется гетерогаметным. Это самцы у млекопитающих и самки у птиц и насекомых.
Половые хромосомы неоднородны по своему генному составу. В процессе изучения геномов млекопитающих было установлено, что у большинства видов, в том числе собаки, человека и других, весьма далеких друг от друга, X-хромосомы имеют идентичный состав и аналогичную морфологию. В Х-хромосоме, кроме генов, обуславливающих развитие пола, присутствуют и гены, определяющие признаки, не относящиеся к нему, например, свертываемость крови и т. д.
В Y-хромосоме локализованы в основном гены, непосредственно относящиеся к полу. Других генов — единицы. Поэтому Y-хромосому часто называют генетически пустой.
Гормональные механизмы определения пола
Начальные этапы развития половых желез одинаковы у самцов и самок. На раннем периоде эмбриогенеза, вскоре после имплантации, из эктодермы возникают зародышевые половые клетки, так называемые гоноциты. Посредством амебоидных движений они передвигаются к зачаткам половых желез, возникшим из мезодермы. В этих зачатках возникают первичные половые тяжи и вскоре зачаточные половые железы оказываются состоящими уже из двух слоев — мозгового и коркового. Такие половые железы способны дифференцироваться как в мужские, так и в женские половые железы.
Дальнейшая эмбриональная дифференцировка определяется генетическим полом зародыша. Гормоны, образующиеся в клетках зачаточных половых желез, обеспечивают дальнейшее развитие половых органов по мужскому или женскому типу. Из первичных половых тяжей образуются семенные канальцы и будущие клетки Сертоли, затем появляются клетки Лейдига, которые и начинают выделять мужской половой гормон тестостерон, относящийся к группе андрогенов. Тестостерон направляет развитие половых органов по мужскому типу. В генетически мужских организмах[1] мозговое вещество зачатков половых желез под воздействием тестостерона дифференцируется в семенники, корковое же вещество по мере развития семенников атрофируется и исчезает. Высокая концентрация андрогенов в эмбриональный период формирует контрольную функцию гипоталамуса над гонадотропной функцией гипофиза: гипоталамус обеспечивает постоянную секрецию гонадотропинов взрослого организма по ацикличному типу, то есть по мужскому типу.
В случае если у генетически мужских особей выработка андрогенов не наступает во время, то развитие половых желез идет по женскому типу, система гипоталамической регуляции гипофиза формируется также по женскому типу (циклично) и вследствие всего вышеописанного развивается зародыш с женскими половыми железами, то есть возникает мужской псевдогермафродитизм или иначе тестикулярная феминизация.
После завершения формирования зародыша активность мужских половых желез снижается, образование андрогенов практически прекращается вплоть до периода полового созревания. В период полового созревания количество вырабатываемых половыми железами андрогенов возрастает, что приводит к развитию вторичных половых признаков и полового поведения. Под влиянием андрогенов интенсифицируется синтез белка, рост костей вначале усиливается, а затем прекращается. Недостаток андрогенов в этот период приводит к продолжительному росту костей и развитию гипогонадного гигантизма.
В генетически женском организме[2] зачатки половых желез в виде тяжей мезотелия, врастающие в эмбриональную закладку яичника превращаются в первичные фолликулы, и дальнейшее развитие половых желез идет по женскому типу: из коркового слоя развиваются яичники, а мозговой слой атрофируется. Система регуляции гонадотропной функции гипофиза формируется также по женскому типу.
В связи с описанной общностью эмбриональных закладок половых органов у самок всегда вырабатывается некоторое количество мужских половых гормонов (андрогенов), а у самцов — женских половых гормонов (эстрогенов).
После рождения животных органы размножения как самцов, так и самок претерпевают важные структурно-функциональные изменения. Так, в первые 6 месяцев постнатальной жизни, абсолютная масса семенников увеличивается в 16–17 раз, а относительная с 0,19 до 0,80% к массе тела. Увеличивается диаметр извитых канальцев, изменяется их клеточный состав. Нарастает масса придаточных половых желез, особенно в период полового созревания.
Половой диморфизм
Признаки, по которым отличаются особи разных полов делятся на первичные и вторичные. Первичные половые признаки это те, которые обеспечивают образование гамет и соединение их в процессе оплодотворения. У всех млекопитающих, в том числе собак, это половые железы (гонады), половые пути и наружные половые органы (гениталии). К вторичным половым признакам относят признаки и свойства организма, не обеспечивающие непосредственно процессы образования половых клеток, спаривания и оплодотворения, но играющие важную роль в половом размножении.
У собак к ним относятся особенности телосложения и поведения, обеспечивающие, так называемый, половой диморфизм. У большинства видов животных половой диморфизм заключается в том, что самцы имеют более развитые вторичные половые признаки по сравнению с самками. До половой зрелости, а точнее до первого сезона размножения, самки и самцы многих видов животных очень похожи друг на друга. Под влиянием половых гормонов внешний вид самцов, а у некоторых видов — самок, заметно преображается. Некоторые рыбы приобретают необыкновенно яркую окраску, у самцов копытных отрастают рога, у некоторых обезьян — гривы, усы и борода. У птиц образуются совершенно невероятные наряды из перьев, отрастают гребни, набухают сережки. У некоторых видов эти изменения сохраняются в течение всей жизни, у других же подобные наряды служат признаки готовности к размножению и проявляются только в брачный сезон. Подобные ритуальные органы обнаруживаются у представителей любой другой группы животного мира. Таковы, в частности, броские, яркие отметины и экстравагантные, удлиненные, расширенные или причудливо вырезанные перья многих птиц, видоизмененные плавники рыб, меняющие окраску кожные «воротники» рептилий. Все эти «украшения» явно демонстрируются перед прочими особями своего вида, перед самкой или соперником за счет специфических форм демонстративного поведения. С приближением сезона размножения под воздействием половых гормонов, животные начинают демонстрировать свои половые признаки. Они поднимают и опускают хохлы, распускают хвосты, как, например, павлины, производят множество ритуальных движений, в общих чертах весьма похожих у представителей разных таксономических групп. Очевидно, в ходе эволюции и сами украшения, и способы их показа развивались параллельно. Демонстрирование этих сигнальных структур несет жизненно важную информацию, которая указывает другим особям на половую принадлежность демонстрирующего животного, на его возраст, силу, право собственности на данный участок местности и т. д.
У собак специфические вторичные половые признаки — как, например усы и борода у человека или хвост у павлина отсутствуют, однако при нормальном развитии самка и самец одной породы имеют выраженные внешние отличия. Половой диморфизм у собак хорошо выражен в специфическом для пола развитии вторичных половых признаков, как, например размеров тела, соотношения мышечной и костной массы, формы головы. Кобели всех пород крупнее и массивнее сук, имеют мужественное выражение морды. Они агрессивнее и сильнее сук. Половому диморфизму придается большое значение в оценке экстерьера собак, несоответствие половому типу относится к разряду дисквалифицирующих пороков. Племенное использование таких собак ведет к появлению разнообразных нарушений физиологических процессов и экстерьера собак.
ГЛАВА 3 ОБРАЗОВАНИЕ ПОЛОВЫХ КЛЕТОК
Размножение становится возможным лишь по достижении животным половой зрелости. Половая зрелость животных наступает с момента начала активного образования половых клеток. Сроки наступления ее у собак сильно варьируют и зависят от многих внешних и внутренних причин.
В процессе с образования половых клеток проходит мейотическое деление клетки, приводящее к уменьшению числа хромосом вдвое. Мейоз обеспечивает сохранение постоянства числа хромосом. Если бы число хромосом в гаметах оставалось таким же (диплоидным) как и в соматических клетках (клетках тела), то при оплодотворении в каждом поколении число хромосом бы удваивалось. Кроме того, в процессе мейоза, каждая из хромосом имеет равную вероятность попасть в ту или другую из образующихся гамет. Соответственно в каждой из гамет могут оказаться как «отцовские», так и «материнские» хромосомы. При диплоидном наборе хромосом у собаки, равным 78, число возможных сочетаний хромосом составит 3003! Вероятность того, что в одну клетку попадут хромосомы только отца или только матери ничтожно мала и составит в данном случае (1/2) в тридцать девятой степени! Дополнительное перемешивание генетического материала осуществляется также путем кроссинговера — обмена участками хромосом в процессе мейоза.
Сперматогенез
Сперматогенез — образование сперматозоидов в семеннике, продолжающееся непрерывно в течение всего периода половой зрелости кобеля. Это сложный и довольно длительный процесс: периоды размножения клеток сменяются периодами роста, созревания, формирования.

Рис. 1. Схема сперматогенеза
Развитие сперматозоидов происходит в семенных канальцах — многочисленных тонких трубочках, из которых состоит семенник. Стенка семенного канальца состоит из тонкой соединительнотканной основы и внутреннего спермиогенного слоя, образованного эпителием, называемого сертолиевым, и располагающимися в его толще мужскими половыми клетками на разных стадиях их развития. Сертолиевы клетки заполняют промежутки между округлыми половыми клетками и образуют ту непосредственную среду в которой происходит развитие последних. Они обеспечивают снабжение половых элементов питательными веществами, приносимыми кровью и осуществляют гуморальное воздействие на половые клетки со стороны организма.
Периоды сперматогенеза
Первый период сперматогенеза — период размножения первичных половых клеток, дающих начало сперматогониям — мелким округлым клеткам, с очень незначительным количеством цитоплазмы в виде тонкой каемки вокруг ядра, энергично размножающихся митозом.
Второй период сперматогенеза — период роста — характеризуется прекращением размножения сперматогониев и превращением их в сперматоциты 1 порядка. Сперматоциты растут, увеличиваясь в размерах в 4 и более раз.
Третий период сперматогенеза носит название периода созревания. Созревание заключается в двух быстро следующих друг за другом делениях сперматоцитов I порядка в результате чего сначала получаются два сперматоцита II порядка, а затем четыре сперматиды.
Это деление представляет собой митоз и образующиеся сперматиды оказываются носителями гаплоидного набора хромосом.
Четвертый период — период формирования спермиев. В течение этого периода сперматиды приобретают специальные приспособления, необходимые для обеспечения процесса оплодотворения и превращаются в сперматозоиды.
Строение сперматозоида
Каждый спермий содержит ядро с гаплоидным набором хромосом, двигательную систему, обеспечивающую движение сперматозоидов и полость, заполненную ферментами, необходимыми для проникновения ядра спермия в яйцо. В процессе созревания ядро сперматиды удлиняется, уплощается и образует основную массу головки сперматозоида. Головка сперматозоида является аппаратом передачи генетической информации, закодированной в ДНК. Значительная часть цитоплазмы элиминируется при созревании спермия и незначительная ее доля сохраняется в виде тонкого слоя вокруг ядра, образуя на переднем конце головки чехлик с заостренным концом. Чехлик содержит в себе акросомный пузырек, образовавшийся из элементов аппарата Гольджи и содержащий ферменты, разрушающие белки и полисахариды оболочки яйцеклетки при оплодотворении. Акросомный пузырек можно, таким образом, рассматривать как видоизмененную лизосому.

Рис. 2. Строение сперматозоида (схема строения спермия)
1 — акросома; 2 — ядро; 3 — шейка; 4 — средний отдел с митохондриями; 5 — хвостик
Самая короткая часть спермия это шейка или иначе — основание ядра.
Средний отдел и хвостик сперматозоида представляют собой его двигательный аппарат. Спермий способен передвигаться при помощи движения жгутиков, имеющих весьма сложное строение. Центрально расположенная аксонема окруженная наружными волокнами, образована девятью периферическими и одной центральной группой микротрубочек, прикрепленных к основанию ядра спермия. Белок динеин, способен преобразовывать химическую энергию, выделяющуюся при расщеплении молекул АТФ, в механическую и, таким образом, приводить жгутики в движение. Энергию для движения жгутиков поставляют в виде АТФ в митохондрии, лежащие в основании ядра спермия, а также митохондрии расположенные в виде спирали вокруг тела спермия. Хвостик сперматозоида подвижен и его движения способствуют активному передвижению спермия. Скорость передвижения спермия в типичных условиях составляет около 2,5 мм в минуту.
Таким образом, сперматозоид является весьма специализированной клеткой, снабженной всеми необходимыми функциональными и морфологическими приспособлениями, обеспечивающими оплодотворение.
Оогенез
Процесс развития женских половых клеток оогенез имеет ряд общих черт с процессом сперматогенеза, характеризуясь, как и последний, наличием периодов размножения оогониев, роста и созревания. Как и при сперматогенезе, созревание женских половых клеток происходит в форме двух последовательных делений созревания и сопровождается уменьшением числа хромосом вдвое с образованием гаплоидных половых клеток — гамет. В то же время процесс оогенеза имеет и существенные отличия по сравнению с развитием мужских половых клеток. Если при сперматогенезе период роста выражен сравнительно слабо, а период формирования характеризуется наиболее существенными преобразованиями развивающейся семенной клетки, то при оогенезе, напротив, гораздо более длительным является период роста, тогда как период формирования отсутствует.

Рис. 3. Схема оогенеза
Периоды оогенеза
Период размножения оогониев проходит в эмбриональном периоде, и самки рождаются уже с ооцитами I порядка, которые, однако, вырастают весьма незначительно и сохраняются в таком состоянии в яичнике до периода половой зрелости организма. С наступлением половой зрелости ооциты I порядка вступают в процесс дальнейшего роста и созревания.
Период созревания заключается, как и при сперматогенезе, в двух последовательных делениях ооцита, приводящих к образованию четырех клеток с гаплоидным набором хромосом. Однако из этих четырех клеток лишь одна превращается в яйцеклетку, три других, так называемые направительные тельца, постепенно редуцируются.
Биологические особенности половых клеток
При всей биологической «равноправности» мужской и женской половых клеток в процессе оплодотворения, в формировании наследственности дочернего организма, перед ними все же стоят совершенно разные функциональные задачи. Яйцеклетка должна обеспечить зародыш будущего организма основной исходной массой цитоплазмы и достаточным количеством питательного материала. Именно поэтому она должна быть сравнительно крупной и не может быть активно подвижной. Яйцеклетка собаки, так же как и всех животных в сотни раз крупнее сперматозоида, ее максимальный диаметр равняется 140 мкм. Вместе с тем, она не способна к самостоятельному делению.
Сперматозоид, напротив, должен обладать активной подвижностью, чтобы быть способным достичь яйцеклетки, и в связи с этим должен быть максимально разгружен от запасных питательных веществ и обладать малыми размерами и значительными запасами энергии. Кроме того, спермии значительно многочисленнее яйцеклеток, так как шанс достичь яйцеклетки для каждого данного спермия невелик. Так у собаки в каждой одновременно выбрасываемой при половом акте порции семени (эякуляте) содержатся сотни миллионов сперматозоидов, а в оплодотворении из них участвуют одни или немногие. Ряд физиологических особенностей спермиев (свойство двигаться против тока жидкости, выделение ферментов определенным образом влияющих на яйцеклетку и т. д.), а также специфических морфологических черт представляют собой весьма совершенные приспособления, обеспечивающие возможность оплодотворения.
ГЛАВА 4 ПОЛОВЫЕ ЦИКЛЫ
Процесс размножения носит циклический характер, т. е. связано с периодическим повторением комплекса специфических явлений. В течение полового цикла в организме животного происходит ряд изменений. Наибольшей глубины эти изменения в период размножения достигают у самки. Половой цикл должен закончиться оплодотворением, если же этого не произошло, самка остается холостой. Половые циклы повторяются до тех пор, пока самка способна к размножению. По достижении определенного возраста эта способность утрачивается, наступает так называемый климактерический период.
По количеству половых циклов различают виды полициклические, у которых половые циклы у самок повторяются несколько раз в году, и моноциклические, имеющие один цикл в году, приуроченный к определенному сезону.
Половая циклика у самцов выражена несколько слабее. Она заключается в основном в изменениях интенсивности сперматогенеза, а также поведения животных.
Половая система высоко чувствительна к изменениям внешней среды. Как показали исследования, на ритм половых циклов оказывает влияние множество факторов, в частности, пища, температура, свет и т. д.
Весь круг явлений, связанный с процессом размножения у зверей в естественных условиях, складывается из нескольких этапов: 1) подготовка к размножению, 2) соединение полов и совокупление, 3) беременность, 4) роды, 5) подсосный период и воспитание потомства, 6) распад семей и расселение молодых. Примерно тот же цикл мы наблюдаем и у собак.
Подготовка к размножению
По мере приближения срока размножения, стадия покоя сменяется стадией возбуждения. Эта стадия характеризуется рядом изменений в половом аппарате самки и общим возбужденным ее состоянием. Под влиянием усиленного продуцирования передней долей гипофиза гонадотропного гормона, в яичниках происходит интенсивное развитие фолликулов. Созревающие фолликулы выделяют в кровь самки гормоны, которые вызывают гиперемию половых органов, увеличение размеров яичников и более или менее ярко выраженное влечение к самцу.
Одновременно с изменениями, происходящими в период течки в половых органах самок, соответствующим образом изменяется и половая сфера самцов. Подготовка к размножению у них заключается в основном в интенсивном созревании и формировании мужских половых клеток — сперматозоидов. Подобно яичникам, семенники и их придатки в период подготовки к размножению также сильно увеличиваются в размерах. У самцов домашних животных, способных к размножению в течение круглого года сезонные изменения в семенниках и минимальны. Однако у собак живущих в условиях приближенных к естественным, например, у северных ездовых или охотничьих собак, сезонность размножения имеет место. У кобелей этих пород, подобно диким животным, наблюдается снижение функции семенников вне сезона размножения. Вполне возможен подобный процесс и у кобелей пород, находящихся в период становления от аборигенной к заводской, особенно при их вольерном содержании.
Вся совокупность сигналов исходящих от другого пола возбуждающе действует на партнеров. У многих видов зверей пришедшие в охоту самки начинают искать сближения с самцами, заигрывают с ними, производят ряд характерных движений, учащают мочеиспускание, перед самым совокуплением принимают позу, облегчающую этот акт. У других видов самки менее активны и внешне остаются спокойными. Самцы, движимые половым инстинктом, начинают настойчиво преследовать самок, гоняются за ними. В связи с этим эту фазу часто называют «гоном».
Таким образом, гон, или брачный период — это комплекс действий, направленных на соединение разнополых особей, т. е. на спаривание, в результате которого совершается акт совокупления.
Половой цикл суки
Строение яичников и других частей полового аппарата суки подвержено более или менее правильным периодическим изменениям, совокупность которых носит название половых циклов. Одновременно с изменениями в половой системе в организме суки происходят определенные ритмические изменения обмена веществ, температуры тела, функционального состояния центральной нервной системы и почти всех внутренних органов. Состояние яичников сказывается на всем организме самки и оказывает влияние на все его функции. Эти ритмические изменения оказываются синхронными с фазами фолликулярного цикла яичников, то есть с ритмическим повторением процессов роста и созревания фолликулов, овуляции и образования желтых тел.
Один из важных признаков, отличающий диких животных от домашних, — моноцикличность, то есть размножение раз в год, в строго определенный, наиболее благоприятный для этого сезон. При таком типе размножения половые железы, как самок, так и самцов функционируют только в сезон размножения. Для домашних животных характерно либо отсутствие приуроченности рождения детенышей к строго определенному сезону, либо возникновение полицикличности, возможности размножения два и более раз в году. У кобелей заводских пород семенники функционируют круглый год.
Самки домашних собак, как правило, имеют два-три половых цикла в течение года, отделенные друг от друга периодами покоя, без видимой привязанности к сезону. Циклика, при этом имеет значительную индивидуальноую или семейную, генетически обусловленную вариабельность от четырех до одного раза в год. В среднем продолжительность полового цикла от начала одной пустовки до начала другой составляет 5–8 месяцев, но возможны породные и индивидуальные колебания от 4 до 13 месяцев. У представителей примитивных и аборигенных пород цикл несколько удлинен, почти до одного раза в год, как у диких представителей псовых и строго приурочен к определенному сезону.
При воздействии различных стрессогенных факторов могут возникать разнообразные отклонения от нормальномого полового цикла, вплоть до полного подавления эструса.
Половая зрелость у сук наступает в возрасте 6–10 месяцев. Вопреки распространенному мнению наступление половой зрелости не зависит от размера и конституции собаки, а определяется, скорее особенностями индивидуального развития. Время начала первой течки даже у однопометных сук может сильно отличаться.
Фазы полового цикла
Половой цикл можно разделить на четыре фазы.
Фаза 1. Проэструс или предтечка
По мнению различных авторов, этот период длится от 3 до 16 дней, в среднем 7–10. В это время в яичниках происходит рост и созревание фолликулов, выделяющих женские половые гормоны — эстрогены. Возрастание их концентрации в крови вызывает прилив крови к родовым путям и матке. Вследствие этого происходит усиление функций слизистых внешних и внутренних половых органов суки. Набухает петля, увеличиваются размеры рогов матки. Стенки матки и влагалища утолщаются, количество кровеносных сосудов в них увеличивается, и начинаются серозно-кровянистые выделения из вульвы.
Мазок, взятый в этот период из влагалища, состоит почти исключительно из эпителиальных клеток, имеющих крупное ядро и расположенных поодиночке или небольшими кучками. Лейкоциты в мазке отсутствуют. Возникает целый комплекс поведенческих реакций: сука становится игривой, непослушной, резко увеличивается мечение территории мочой; но попытки кобелей сделать садку в этот период вызывают резкую агрессию со стороны суки.
Проэструс продолжается в среднем 7–9 дней, но может варьировать от 3 до 18 дней. Кровянистые и слизистые выделения из петли также могут иметь разную интенсивность. Иногда у сук бывает так называемая «сухая» течка, практически без выделений, что, однако, не препятствует нормальному созреванию фолликулов, оплодотворению и эмбриональному развитию плодов.
Неопытные кобели остро реагируют на суку во все периоды течки, делая попытки спаривания. Опытные кобели обычно не делают никаких попыток вне периода охоты. Однако если сука вызывает реакцию кобелей практически постоянно — вероятнее всего по какой-либо причине (воспалительные, или онкологические заболевания половых органов) у нее нарушен гормональный статус.
Фаза 2. Собственно эструс — «охота»
Длится от 4 до 12 дней. Видимые выделения из петли в это время становятся менее кровянистыми и приобретают светловатый оттенок.
В этот период в яичниках происходит быстрый предовуляционный рост наиболее крупных пузырчатых фолликулов и их овуляция (разрыв). Поверхностные слои эпителия влагалища ороговевают и слущиваются. Влагалищный мазок состоит только из ороговевших безъядерных клеток, имеющих вид чешуек. Лейкоциты и эпителиальные клетки с ядрами в мазке совершенно отсутствуют. В конце течки ороговевшие чешуйки образуют скопления.
В эту фазу эстрального цикла сука готова к спариванию. В половых путях суки накапливается большое количество ферментов, повышающих эластичность тканей и способствующих жизнеспособности, продвижению и оплодотворяющей способности спермиев. Петля резко увеличивается в размерах и визуально становится более заметной, наощупь кажется мягкой и податливой. Изменяются поведенческие реакции суки. При прикосновении к крупу, и, тем более, петле, сука поддергивает петлю вверх, отводит хвост в сторону и замирает в стойке. При приближении кобеля принимает позу готовности к спариванию. Этот период характеризуется изменением поведения суки — она становится игривой, беспокойной. Овуляция происходит как правило на 1–3-й день охоты. Все созревшие фолликулы овулируют в течение 12–48 часов. Яйцеклетки попадают в яйцеводы и движутся по направлению к матке. Однако яйцеклетки у собак выходят из фолликулов еще незрелыми. Они приобретают способность к оплодотворению только после 3-х дневного пребывания в яйцепроводе и матке и затем сохраняют ее в течение суток. Таким образом, в большинстве случаев, оптимальным временем вязки считается 2–4 день от начала охоты. Так как выход яйцеклеток происходит неодновременно, то для лучшей завязываемости плодов — желательно проведение контрольной вязки через 24–48 часов. В половых путях суки сперматозоиды сохраняют оплодотворяющую способность до недели, в среднем — около полутора суток. Сам процесс оплодотворения — сложная физико-химическая реакция с участием ферментов и антител. На всех трех стадиях — продвижения спермиев по половым путям до яйцеклеток, оплодотворения и имплантации зародышей (прикрепление к стенке рога матки) — происходит мощный конкурентный отбор, и от его степени зависит качество будущего потомства.
Фаза 3. Метаэструс — период «после течки»
Лютеональная фаза. После овуляции на месте лопнувших фолликулов образуются так называемые желтые тела, продуцирующие гормон прогестерон, который стимулирует секреторную функцию эндометрия, создавая благоприятные условия для имплантации оплодотворенных яйцеклеток и развития беременности. С наступлением метаэструса самка перестает допускать к себе кобелей, постепенно исчезает отечность вульвы и прекращаются выделения. Эпителиальный слой эндометрия претерпевает перерождение. Лейкоциты инфильтруют стенки матки и влагалища. Влагалищный мазок содержит немало ороговевших чешуек и много лейкоцитов, на короткое время в нем могут появляться также эпителиальные клетки с ядрами. Секреция прогестерона желтым телом достигает максимума к 20–30-му дню метаэструса, независимо от того, наступила беременность или нет, а затем к 70–80-му дню медленно снижается, при наличии щенности — к 60–65-му дню, то есть к началу родов.
В этой стадии цикла у сук нередко возникают заболевания матки, в частности пиометра, чему способствуют продолжительные, повторяющиеся при каждом очередном цикле воздействия прогестерона на матку, предрасполагающие к развитию гиперпластических и кистозных изменений в эндометрии и повышающие его чувствительность к инфицированию.
Под влиянием гормона желтого тела в обоих рогах матки развиваются секреторные изменения, мало отличающиеся от наступающих во время беременности. Такое состояние самки называют ложной беременностью.
Фаза 4(0). Анэструс — половой покой
Длится примерно три — четыре месяца. В это время происходит обратное развитие слизистой матки и влагалища, снижение эластичности и уменьшение размеров. Этот период характеризуется уравновешенностью общего состояния организма, необходимой для накопления сил и подготовки к размножению. Однако, «покой», в котором находится в это время организм, относителен: половые железы многих видов продолжают исподволь продуцировать половые клетки. На самом деле такому определению отвечают лишь внешние проявления взаимоотношений между полами.
По окончании брачного периода возбуждение самцов и самок спадает. Если не наступила беременность, то в половом цикле самок наступает стадия торможения, характеризующаяся угасанием полового тонуса, уменьшением яичников. Организм животного, таким образом, снова возвращается к состоянию покоя.
Большинство же особей после периода спаривания приступают к подготовке к выращиванию потомства.
Эндокринологическая картина полового цикла
В период анэструса уровень половых стероидов — прогостерона и эстрадиола в крови очень низок. Примерно за четыре недели до наступления следующей течки отмечается легкое повышение уровня эстрадиола. К началу течки рост фолликулов стимулирует внутреннюю секрецию эстрадиола, содержание которого в конце предтечковой стадии дает отчетливый резкий всплеск.
Начало течки характеризуется резким падением уровня эстрадиола и одновременно медленным ростом уровня прогостерона. В это время происходит овуляция. После быстрого подъема, за одну неделю, уровня прогостерона в начале эструса его количество в течение примерно 3-х недель колеблется на достаточно высоком уровне, примерно в 30 раз превышающем основную величину, и в конце метаэструса снова опускается до основной величины.
В последние годы появился экспресс — тест для прблизительного количественного определения уровня прогостерона в крови. Так как уровеньпрогостерона с достаточной степенью постоянства начинает повышаться за 2–3 дня до овуляции, то предполагалось, что по результатам этого теста возможно довольно точное определение оптимального срока вязки. Однако сравнительные исследования с помощью обычных гормональных методов показали, что этот тест во многих случаях неточен.
Определение оптимального срока вязки
Вязка суки оказывается наиболее результативной, если она происходит в момент овуляции. К сожелению, метода, позволяющего абсолютно точно определить время овуляции, пока не существует. При определении оптимальных сроков вязки используются опосредованные методы, ориентирующиеся либо на изменения структуры влагалища под действием повышенного количества эстрогенов во время предтечковой фазы, либо на повышение уровня прогостерона непосредственно перед овуляцией. Однако, у разных сук возможны значительные колебания в этих процессах, поэтому ни одна из диагностических методик не является абсолютно надежной. Исходя из этого, окончательный диагноз предпочтительно ставить на основании результатов различных исследований.
Вагинально-цитологическая диагностика
Гистологическое строение влагалищного эпителия подвержено гормональным влияниям. В анэструсе влагалище покрыто 2–3-слойным неороговевшим плоским эпителием. Под влиянием эстрогенов происходит пролиферация клеточных слоев, и клетки, находящиеся на поверхности, подвергаются ороговению. Пик пролиферации (до 20 слоев и ороговение, как правило, совпадают с течкой. Вначале метаэструса эпителий опять постепенно уменьшается.
При вагинально — цитологическом обследовании это гормонально обусловленное изменение структуры используется для определения фазы цикла, оптимального срока вязки, и рапознавания дисфункции яичников.
В процессе исследования с помощью влагалищного тампона берется клеточный материал из верхних слоев эпителия. При наличии течки и готовности к вязке в мазке будут преобладать ороговевшие эпителиальные клетки, в фазе анэструса будут обнаруживаться только неороговевшие клетки. Проведение вязки рекомендуется при ороговении более 80% суперфициальных клеток. Максимальное количество ороговевших клеток сохраняется 1–6 дней. В конце течки эпителиальный слой в течение нескольких дней отшелушивается и ороговевшие клетки в мазке встречаются в соединениях. В этом процессе регенегации участвуют также лейкоциты, которые юывают представлены в большом количестве в мазках, полученных между 2 и 4 днями метаэструса.
Максимальное ороговение эпителия влагалища у 75 процентов сук приходится на время овуляции, в исключительных случаях за 10 дней до ее начала или через день после овуляции. В связи с этим, если у суки наблюдаются проблемы с зачатием, одного цитологического метода может быть не достаточно.

Рис. 4. Схематическое изображение различных эпителиальных клеток влагалища в фазах эструса и анэструса
Отклонения от нормального протекания полового цикла у собак
Нормальный половой цикл у суки подвержен довольно значительной изменчивости. Е.Е. Коваленко (1995) на основании исследования большого количества сук приходит к выводу о том, что длительность отдельных фаз полового цикла может варьировать независимо друг от друга. Фаза эструса может иметь два или больше пиков овуляции. В связи с этим, на протяжении такого эструса готовность суки к спариванию не всегда одинакова. Автор приводит ряд вариантов протекания фаз овуляторного цикла представляющий собой переходы от наиболее обычного варианта к крайним отклонениям.
Наблюдающиеся у сук отклонения от нормального хода полового цикла, вызваны нарушениями гормональной регуляции, которые, в свою очередь, могут быть связаны с аномалиями строения соответствующих желез, воспалительными процессами и воздействиями окружающей среды.
Анэстрия
Анэстрия — отсутствие течки. Е.Е. Коваленко (1993) выделяет следующие типы анэстрии: врожденную, приобретенную, постоянную и временную, анэстрию полового созревания (отсутствие течки у суки до 14–18 месяцев), юношескую (отсутствие течки после 1–2 нормальных течек), послеродовую (отсутствие течки после родов), скачущую (периодическое увеличение периода анэструса более, чем на 10 месяцев).
Причиной этих отклонений может служить как недоразвитие яичников и, как следствие этого, низкая концентрация эстрогенов в крови; так и недостаточность гонадотропных гормонов гипофиза (лютеинизирующего и фолликулостимулирующего) и щитовидной железы (тиреотропного), а также гормонов половых желез — эстрогенов. Истинную причину анэстрии позволяет выявить исследование концентрации гормонов в крови. Врожденная анэстрия развившаяся вследствие недоразвития яичников сопровождается недоразвитием молочных желез и вульвы, что значительно облегчает постановку диагноза. В зависимости от степени недоразвитости яичников течка может отсутствовать полностью или быть скрытой.
Субэстрия
Субэстрия — атипичная течка, в ходе которой трудно выявить отдельные стадии, наступившая в срок. Период охоты в этом случае оказывается смазанным.
Пролонгированная (продолжительная) течка
При пролонгированной течке продолжительность всех или отдельных фаз увеличена (иногда до нескольких месяцев). Понятие «пролонгированная течка» используется весьма широко. Обычно это явление связано с гиперфункцией яичников и часто является следствием кисты яичников, продуцирующей большое количество эстрогенов. Основные отклонения, наблюдаемые при этом можно разделить на три группы: затяжной проэструс, затяжной эструс и расщепленную течку.
Затяжной проэструс — кровянистые выделения, продолжающиеся более 21 дня и не сопровождающиеся признаками готовности суки к вязке. Как предполагают Ниманд и Суттер (1998), причина этого явления заключается в недостаточности выделения гонадотропноых гормонов.
Затяжной эструс — возникает на фоне излишне длительного пика выделения эстрогенов, однако, не приводящего к овуляции. При этом наблюдаются сукровичные выделения и готовность суки к вязке в течение более трех недель. Затяжное действие эстрогенов в этом случае часто приводит к развитию железисто-кистозной гиперплазии эндометрия.
Расщепленная или скрытая течка характеризуется наличием двух волн. В первой волне обнаруживается отек вульвы, выделение крови из влагалища и запах, привлекающий кобелей. Затем течка прекращается вследствие недостаточного синтеза одного из рилизинг-гормонов гипофиза — лютеинизирующего гормона стимулирующего созревание фолликулов. Вязки чаще всего не происходит или при наличии вязки не происходит оплодотворения, сука остается пустой. Во второй волне, наступающей через 2–12 недель, прохождение фаз и поведение суки типично для нормальной течки. Вязка проведенная в этой фазе, как правило, приводит к оплодотворению. Расщепленная течка не может считаться патологической для сук в возрасте до двух лет (с неустановившимся половым циклом).
Воспалительные заболевания матки иногда могут симулировать пролонгированную течку. В обоих случаях суке требуется квалифицированное лечение.
Методы стимулирования и предотвращения течки
При возникновении необходимости стимулирования течки можно использовать введение экзогенных эстрогенов и простагландинов. Вязку или искусственное оплодотворение при этом рекомендуется проводить под контролем влагалищных мазков.
Существует также ряд гормональных препаратов, применяемых с целью супрессии течки.
Однако следует учитывать, что любое вмешательство в ход нормальных гормональных процессов может привести к нежелательным последствиям и должно проводится только под руководством грамотных специалистов.
Кастрация
В последние годы эта процедура приобретает все большую и большую популярность как за рубежом, так и в России. Кастрация — оперативное удаление половых желез. Вследствие возникновения глубоких гормональных нарушений — резкого снижения уровня андрогенов у кобелей (за исключением незначительного количества синтезируемого надпочечниками) и эстрогенов у сук возникает пожизненное бесплодие. В результате изменяется и поведение животных. Исчезают или снижаются проявления половой активности, течки, прекращается или значительно снижается мечение территории, снижается агрессивность. При ранней кастрации (до установления общего гормонального фона и возникновения полового поведения) снижается общая активность животного и как следствие нарастает вес животного.
Кастрация кобелей применяется как с целью ликвидации полового поведения (например, кастрация бродячих животных, широко практикуемая во всем мире), так и с лечебными целями при воспалительных процессах в семенниках, опухолях семенников склонных к малигнизации и злокачественных опухолях. В некоторых случаях с целью предупреждения злокачественного перерождения неопустившегося семенника производят кастрацию крипторхов. Показаниями к кастрации могут служить и поражения мошонки.
Кастрация сук проводится по аналогичным показаниям, включая злокачественные новообразования, в том числе молочных желез.
Стерилизация
Стерилизация животных применяется значительно реже. При этой операции сохраняют половые железы и накладывают лигатуры на семенные канатики у самцов и яйцепроводы у самок, что предупреждает оплодотворение при сохранении гормонального статуса и поведения. Способность к вязке при этом сохраняется.
В настоящее время стерилизация и кастрация неплеменных животных обоих полов приобретает все большую и большую популярность. Во многих странах, например США и развитых странах Европы, кастрируют практически всех пользовательных собак и «домашних любимцев», а также собак, вышедших из племенного возраста. Ветеринарные специалисты часто используют кастрацию сук для предупреждения развития разнообразных дисфункций половой системы и малигнизации предопухолевых заболеваний. Однако, необходимо иметь в виду, что в ряде случаев, снижение уровня половых гормонов в крови кастрированной собаки может иметь и целый ряд побочных действий. Подобным осложнениям особенно подвержены крупные собаки.
«Противозачаточные» средства
Подавление синтеза фолликулостимулирующего гормона при помощи гормональных фармпрепаратов может быть использовано для отодвигания времени начала или прерывания начавшейся течки. Следует учитывать, что дача подобных препаратов суке повышает риск заболевания пиометрой и способствуает аномалиям развития уже завязавшихся плодов.
Препараты мужских половых гормонов, предотвращающие овуляцию, оказывают влияние на действие лютеинизирующего гормона и способны прервать начавшуюся течку. Однако одним из факторов риска при этом может быть маскулинизация суки и нарастание ее агрессивности.
ГЛАВА 5 ПОЛОВАЯ СИСТЕМА СОБАКИ
Половые органы в процессе функционирования создают предпосылки к продлению жизни не отдельного организма, а животного как вида. Наряду со своей основной функцией — размножением, половые органы синтезируют гормоны, влияющие на развитие и жизнедеятельность организма, и таким образом представляют собой железы внутренней секреции[3].
Половые органы самца и самки имеют как целый ряд общих черт, так и ряд специфических отличий.
В половых железах самца — семенниках в огромном количестве развиваются мелкие, снабженные жгутиками, подвижные половые клетки — спермии, а в половых железах самки — яичниках — сравнительно немногочисленные, довольно крупные, округлые, неподвижные половые клетки — яйцеклетки.
Половая система кобеля
Половые органы самца включают парные семенники с придатками, семяпроводы и семенные канатики, придаточные половые железы, непарные мошонку, мочевой канал, половой член и препуций.
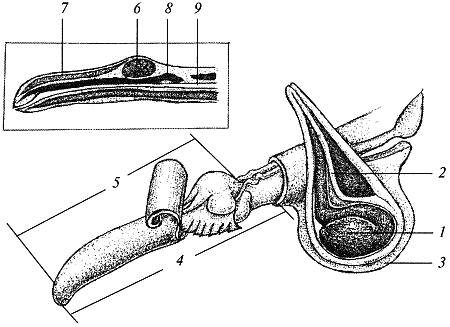
Рис. 5. Половые органы кобеля: 1 — семенник; 2 — семяпрвод; 3 — мошонка; 4 — половой член; 5 — головка полового члена; 6 — пещеристые тела полового члена — «луковицы»; 7 — пещеристое тело длинной части члена; 8 — кость полового члена; 9 — мочевой канал
Семенники
Основными половыми органами самцов являются парные семенники. В них происходит развитие и созревание спермиев. Одновременно семенники представляют собой эндокринные железы и вырабатывают мужские половые гормоны — андрогены.
Строние семенников
Семенники имеют яйцевидную форму. Размеры семенников зависят от размера и веса собаки. У кобеля весом 10 кг их размер составляет примерно 3×2,1×1,9 см., их вес достигает 8 гр.
Каждый семенник подвешен на семенном канатике; с ним тесно связан придаток семенника. Снаружи семенник покрыт плотной и упругой соединительнотканной капсулой, называемой белочной оболочкой. На задней поверхности семенника белочная оболочка образует утолщение, которое носит название средостения яичка. От средостения отходят радиально внутрь плотные соединительнотканные перегородки, делящие паренхиму семенника на конические дольки. Каждая долька содержит семенные канальцы, промежутки между которыми заполнены соединительной тканью. Семенной каналец состоит из длинной извитой части, где происходит сперматогенез, и короткой прямой части, ведущей к средостению и соединяющийся там с соединяющимися друг с другом канальцами сети семенника. Стенки извитой части семенных канальцев содержат расположенные в несколько рядов мужские половые клетки на разных стадиях сперматогенеза, а также так называемые сертолиевые клетки, обеспечивающие питание половых клеток. По мере развития половые клетки продвигаются от наружной поверхности семенных канальцев к их просвету. Когда наступает созревание сперматозоидов, ближайшая к ним часть стенки канальцев расплавляется ферментами и сперматозоиды оказываются в просвете канальца, окруженные небольшим количеством жидкости. В промежутках между семенными канальцами среди нежной соединительной ткани долек расположены группы интерстициальных клеток Лейдига, содержащих зернышки липидов и пигмента. Интерстициальная ткань семенника выделяет мужской половой гормон и относится, следовательно, к эндокринной части этого органа.
Придаток семенника является частью выводного протока. Он служит для накапливания зрелых спермиев, которые могут сохраняться в нем, в течение довольно длительного времени в неподвижном состоянии. Секрет придатков обеспечивает сохранение жизнеспособности и подвижности спермиев. В процессе спаривания животных перистальтические сокращения мышц придатка выбрасывают спермии в семяпровод.
Придаток семенника состоит из трех частей: головки, тела и хвоста. В головке расположены от 7 до 20 семявыносящих канальцев. Эти канальцы начинаются из сети семенника, выходят из его головчатого конца и впадают в проток придатка. Выносящий каналец представляет собой дольку придатка. Диаметр каждого выносящего канальца составляет около 0, 1 0,3 мм. Проток придатка сильно извилист и достигает большой длины. В хвостовой части придатка проток незначительно расширяется, резко поворачивает в направлении головчатого конца и переходит в семяпровод. Хвостовая часть придатка соединена с одной стороны с семенником связкой семенника, а с другой стороны с наружным листком влагалищной оболочки паховой связкой семенника.
Кровообращение семенника осуществляется главным образом конечной ветвью внутренней семенной артерии, проходящей в составе семенного канатика. Ее разветвления проникают внутрь семенника через средостение и образуют в дольках густую капиллярную сеть. Вены идут параллельно артериям. Лимфатические сосуды образуют сеть под белочной оболочкой. Иннервация осуществляется как парасимпатической, так и симпатической нервной системой. Нервные волокна образуют сплетения и проходят внутрь семенника вдоль кровеносных сосудов. Часть волокон проникает в собственную оболочку семенных канальцев, образуя там пуговчатые окончания, или вступает в контакт с интерстициальными клетками.
Мошонка
Семенники вместе с придатками располагаются в кожшом мешке — мошонке. Анатомически мошонка представляет собой выпячивание брюшной стенки, носящего название прямокишечно-пузырного (между прямой кишкой и мочевым пузырем). Таким образом, она образована теми же слоями, что и брюшная полость: висцеральным и париетальным листками серозной влагалищной оболочки и лежащей между ними влагалищной серозной полостью.
Кожа мошонки покрыта мелкими волосками, имеет потовые и сальные железы. По средней линии на ней расположен так называемый шов, мошонки.
Под кожей мошонки находится очень прочно с ней срощенная мышечно-эластическая оболочка. Она развивается из подкожного слоя, поверхностной фасции и гладкой мышечной ткани. Мышечно-эластическая оболочка образует перегородку мошонки, разделяющую ее полость надвое. К мышечно-эластической оболочке прикрепляется подниматель семенника, заключенный в фасцию. Понижение температуры окружающей среды, боль, испуг могоут вызывать сокращения поднимателя и подтягивание семенника к паховому каналу.
Таким образом семенники с их придатки защищены о внешних воздействий следующими слоями: мышечно-эластической оболочкой, фасцией поднимателя семенника, париетальным листком серозной оболочки, влагалищной полостью и висцеральным листком серозной влагалищной оболочки.
Благодаря обилию кровеносных сосудов и потовых желез в коже мошонки температура внутри нее на несколько градусов ниже, чем в брюшной полости. Это способствует нормальному функционированию семенников. Перегрев семенников в брюшной полости в случае задержки их опускания прекращает сперматогенез и ведет к дегенерации семенных канальцев. Перемещение семенников из брюшной полости в мошонку происходит в начале постнатального периода. Задержка одного или обоих семенников в брюшной полости называется крипторхизмом.
Семявыносящий проток, мочеполовой канал
Продолжением протока придатка в виде узкой трубки из слизистой, мышечной и серозной оболочек является семевыносящий проток. Входя в состав семенного канала, семявыводящий проток проходит через паховый канал в брюшную полость, тазовую и паховую области. В тазовой полости семявыносящий проток образует расширенную железистую часть — ампулу и затем открывается семяизвергательным каналом на семенном холмике в начале мочеполового канала или мужской уретры, служащего для выведения мочи и спермы. Этот канал начинается с внутреннего отверстия уретры от шейки мочевого пузыря и оканчивается наружным отверстием уретры на головке полового члена. Только начальная, очень короткая часть уретры — от шейки мочевого пузыря до впадения семяизвергательного канала проводит только мочу, остальная часть — как мочу, так и сперму. Мочеполовой канал состоит из широкой тазовой, проходящей внутри таза, и более узкой половочленной (губчатой) части. В месте перехода от тазовой к половочленной части уретра несколько сужается. В тазовую часть мочеполового канала открываются протоки лежащих рядом придаточных половых желез. Стенка уретры состоит из мышечной оболочки, губчатого слоя и слизистой оболочки. Мышечная оболочка состоит из толстого слоя кольцевых мышц, состоящих из поперечно-полосатой ткани. В тазовой части она образует луковично-кавернозную мышцу, заканчивающуюся на корне полового члена. Губчатый слой имеет основу, состоящую из гладких мышечных и эластических волокон, переплетенных с густой сетью кровеносных сосудов — вен. Особенностями этих вен является наличие специфических расширений — лакун. Густая венозная сеть может заполняться кровью, что приводит к открытию просвета уретры. Слизистая оболочка уретры собрана в продольные складки и в месте впадения семяизвергательных каналов утолщена.
Предстательная железа
Предстательная железа представляет из себя одну из придаточных половых желез, основной функцией, которой является выделение секрета, активизирующего рподвижность сперматозоидов. В зависимости от размеров и возраста кобеля о
Date: 2016-06-06; view: 719; Нарушение авторских прав