Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Ориентировочная реакция и привыкание
|
|
В первой главе были проанализированы два основных класса переменных, характеризующих деятельность мозга. Взятый в качестве модели процесс воспроизведения речи показал возможности, открывающиеся благодаря модификации этих двух переменных. Во второй главе были детально рассмотрены данные, свидетельствующие о пластичности мозга и его способности к постоянным изменениям; однако мы еще не говорили о свойствах самого механизма, который делает возможным осуществление таких изменений. При игре на скрипке, флейте или органе участвуют те же самые основные переменные, что и в речевом общении, хотя сами инструменты различаются тем, как они воспроизводят звуки.
Поэтому настоящая глава начинается с рассмотрения организации нервной системы, позволяющей ей быть аппаратом, создающим все богатство нашего опыта и поведения. Эта организация во многом зависит от относительно постоянных структур, образующихся в тех системах, посредством которых мозг приходит в соприкосновение со своим окружением. Благодаря своей доступности эти системы могут служить некоторыми частными моделями и, следовательно, подвести к исследованию более центральных процессов, анализ которых требует иногда больших усилий. Поскольку довольно сложная организация имеет место уже при первоначальном контакте организма с его окружением, уместно начать наше исследование именно с этого процесса.
Экспериментальные данные говорят о том, что в любой момент времени происходит отбор сенсорного возбуждения с помощью хранящейся в памяти информации о прошлом опыте. Этот процесс сравнения, в ходе которого выявляется, соответствует ли текущее возбуждение хранящимся в мозгу записям, направляет как внимание человека, так и его действия.
Е. Н. Соколов (1960) провел следующий простой опыт, в котором обнаружилось одно из тех противоречий, которые порождают
полезные парадоксы и указывают направление экспериментальным исследованиям в различных разделах нейропсихологии. Человеку предъявляется звук гудка. Сначала он вызывает ориентировочный рефлекс. Электрическая активность мозга характеризуется следующими чертами — активацией в латеральных отделах •больших полушарий головного мозга (в виде увеличения доли низкоамплитудных и высокочастотных компонентов ЭЭГ) и гиперсинхронизацией в медиальных и базальных структурах мозга, на что указывает возрастание ритмичности и рост амплитуды волн среднечастотного диапазона. Кроме того, возникают другие локазатели ориентировочного рефлекса: увеличивается приток крови к голове за счет оттока от кончиков пальцев; меняется электрическое сопротивление кожи, частота сердцебиения и дыха-лия. Однако при многократном повторении звука гудка все эти реакции заметно уменьшаются. В таком случае говорят, что испытуемый обнаружил привыкание к стимулу.
Однако исчезновение реакции на повторяющиеся звуки оказывается обманчивым. В действительности многие из компонентов этой реакции еще сохраняются, так как, если стимул слегка меняется (например, звук становится тише), все первоначально исчезнувшие реакции восстанавливаются. Конечно, это парадокс. До открытия этого явления Соколовым высказывалось предположение, что привыкание просто повышает порог нервной системы на входе. Результаты, полученные Соколовым, означают, что лицо, которое обнаружило привыкание, должно было сопоставить действующий звук с сохранившимся представлением о ранее воспринятых звуках — иначе почему бы снижение интенсивности звука снова вызвало полноценную ориентировочную реакцию? Соколов проверил свою интерпретацию, многократно предъявляя испытуемому тон определенной длительности. Затем он неожиданно предъявлял более короткий звуковой раздражитель. Сразу же после прекращения звука возникали ориентировочные реакции; таким образом, изменение ответов происходило при наступлении тишины. Эти реакции продолжались в течение всей «ожидаемой» длительности звука, а затем медленно исчезали. Мы все испытывали эту удивительную реакцию на неожиданное наступление тишины (рис. III-1). Я назвал это «феноменом Бовери». В течение многих лет на Третьей авеню в Нью-Йорке существовала надземная железная дорога, которая производила ужасающий шум; когда она была снесена, люди, жившие в домах вдоль железной дороги, периодически просыпались от отсутствия звуков во время сна и звонили в полицию, чтобы сообщить, что происходит нечто странное, хотя они не могли точно определить, что именно. Эти звонки раздавались в те моменты, когда раньше с грохотом проносились поезда. Появление чего-то странного было, конечно, связано с наступлением «внезапной тишины», которая заменила ожидаемый шум.
CmiiM.
-щ
ЭЭГ •'(^/йA^^V^ii^Ж^^^
КГР
~Ш~
^/-"'^''Ф**^
КГР
(1В0)
Сттшм.
КГР -
ЭЭГ H№r*^j»4M*«#№A4ll1l^^
В
Сшим КГР
ЭЭГ

^»л^|(Дч^Ч11|^ч,Л^^
Сгтшм.,^^удо^^^^^ Д
ЭЭГ
КГР
зэг
КГР
Рис.III-L Л — угашение ориентировочного рефлекса (ЭЭГ и КГР) на раздражитель интенсивностью, условно обозначенной как 160; Б — растормаживание ориентировочного рефлекса при снижении интенсивности раздражителя до 120; В — то же самое, что и А; Г — возникновение ориентировочного рефлекса на уменьшение длительности раздражителя; Д — то же, что в А; £ — возникновение ориентировочных реакций на неожиданное увеличение длительности раздражителя. Следует отметить, что КГР также возникает с задержкой, что обусловлено замедленностью процессов в автономной нервной системе (Е. Н. Соколов, 1960).
Множество данных свидетельствует о том, что ориентировочные реакции и привыкание длятся сравнительно долго и являются в высшей степени специфическими. Каждая часть мозга характеризуется своими электрическими сигналами и своей последовательностью развития электрических явлений при осуществлении ориентировочной реакции. Если возникает привыкание, то нарушение его (ориентировочная реакция) при всяком изменении ситуации продолжает возникать на протяжении шести месяцев; изменения в последовательности событий, в обстановке, в которой они происходят, или в интенсивности, в длительности или конфигурации самих событий будут вызывать восстановление ориентировочных реакций.
Нейрофизиологии привыкания посвящено много исследований, и она вызвала много споров из-за различных экспериментальных подходов к решению этой проблемы. Для того чтобы стало возможным объяснение этого явления, необходимо уточнить некоторые факторы. Большинство нейрофизиологов называют привыканием любое постепенное ослабление ответа при повторном предъявлении того же самого стимула. Психологи же считают привыканием только те случаи, когда затухание ответа имеет место после нескольких проб и когда легко вызвать восстановление ориентировочной реакции. Недавно некоторые исследователи мозга, в том числе и я, приняли еще более строгий критерий Соколова: неожиданное уменьшение интенсивности, длительности стимула или его отсутствие должны вызывать восстановление ориентировочного рефлекса, если прежде этот рефлекс угасал в результате привыкания.
Важно также место электродов при электрофизиологической записи. Так, в моей лаборатории мы обнаружили, что привыкание действительно характеризуется затуханием ответа на любом уровне зрительной системы обезьяны от сетчатки до коры головного мозга. Однако при разном расположении электродов в коре одни из них дают признаки ослабления, другие — увеличения ответа, а третьи не обнаруживают никаких изменений. Следовательно, при повторении стимула в коре возникает изменение структур возбуждения; растормаживание разрушает эту структуру. Если бы мы смешали записи от всех кортикальных отведений или сделали бы записи от черепа, а не через электроды, вживленные непосредственно в кору, мы не смогли бы обнаружить этот эффект, о чем действительно сообщают некоторые исследователи.
Много работ было выполнено и на клеточном уровне. Была произведена запись от отдельных клеток спинного мозга и других отделов мозга. Во многих местах были обнаружены нервные клетки с затуханием ответа при повторении стимуляции. Такая реакция возникает даже у беспозвоночных, например у Aplesya Cali-fornica (Pinsker et al., 1970; Kupfermann et al., 1970; Castellucci
«et al., 1970). Однако можно ли у всех нейронов, у которых обнаружено привыкание, вызвать явления растормаживания? Недавно полученные Р. Томпсоном данные (Grove, Thompson, 1970) говорят о том, что можно различать по крайней мере три группы нейронов: нейроны, у которых реакция просто затухает; нейроны, реакция которых первоначально усиливается; и нейроны, характеризующиеся сначала усилением, а затем затуханием реакции. -Лишь у последней группы возможно растормаживание. Такие эксперименты до сих пор были проведены только на спинном мозге. Другие отчеты об экспериментах на структурах ствола мозга (верхнее двухолмие) свидетельствуют о том, что возможно, хотя и не обязательно, что одиночные нейроны с затухающими ответами в этой структуре могут обнаруживать растормаживание (Gerbrandt, Bures, Buresova, 1970).
Эти данные, а также другая работа Томпсона (Thompson and Spenser, 1966) показывают, что привыкание связано с взаимодействием нескольких нейронов. Томпсон использовал препарат, принесший известность Шеррингтону: животное, у которого спинной мозг отделен от головного. Повторная стимуляция электрическими залпами входных волокон спинного мозга приводит к постепенному уменьшению рефлекса сгибания конечности (вызванного подобно коленному рефлексу у больного, подвергающегося врачебному осмотру). Этот препарат создавал идеальные условия для изучения места и физиологической природы тех изменений в спинном мозге, которые связаны с привыканием (рис. III-2). Ни входные, ни выходные нейроны не участвуют в изменении рефлекса и, следовательно, не ответственны за привыкание: решающее значение имеют интернейроны, связывающие нейроны входа и выхода. Во-вторых, вопреки ожиданию введение фармакологических веществ, обычно используемых нейрофизиологами при изучении пре- и постсингштических.эффектов, не оказывало никакого влияния; эти вещества не приводили к затуханию ответа, свидетельствующему о привыкании. Это значит, что привыкание не связано с изменениями медленных потенциалов соединительных контактов. Сходные результаты получил Соколов (Соколов, Пакула и Аракелов, 1970). Он обнаружил, что медленные потенциалы мембраны нейрона и генераторные потенциалы, записанные от его аксона, которые дают начало нервным импульсам, могут изменяться независимо друг от друга. Кроме того, полученные им результаты свидетельствуют о том, что механизм генерации нервных импульсов может обнаружить привыкание.
Таким образом, в некоторых случаях процессы затухания, лежащие в основе привыкания, по-видимому, связаны не с пре- ж постсинаптическими изменениями, а с какими-то другими механизмами. Однако в других случаях были получены противо-

Рис. 111-2, Затухающий ответ эфферентной единицы на повторяющееся электрическое раздражение кожи (Thompson, 1967).
положные результаты. Иногда \по не всегда) этот процесс связан с уменьшением или увеличением притока медиаторных веществ,
С. Шарплесс (1967, 1969) показал, что при стимуляции нейронов у позвоночных не возникает ни сенситизации, ни десенси-тизации их сияаптических мембран. Напротив, Э. Кендел (Castel-lucci et al., 1970; Kupfermann et al., 1970; Pinsker et al., 1970) нашел, что привыкание и его нарушение у беспозвоночных локализуются в пресинаптических окончаниях и предполагают участие эффективного механизма секреции возбуждающих веществ в синапсах.
Стало быть, полученные данные не согласуются между собой. Необходимо найти ответ на целый ряд вопросов. Каким образом вообще механизм возникновения возбуждений может испытывать влияния, если не через посредство медленных потенциалов? Запускается ли внутри нейрона некоторый биохимический процесс и если да, то как (особенно если мы имеем дело с системой, которая обычно обнаруживает спонтанную активность и, следовательно, каким-то образом генерирует потенциалы)? Какова природа биохимических изменений и как они
/Р
развиваются во времени? Соколов предполагает, что в этом механизме участвует РНК. Если он прав, то не связано ли привыкание с первыми этапами индукционного процесса или механизм привыкания совершенно независим от процесса постоянного хранения следов памяти? Возможно, что механизм постоянного хранения действует только при условии длительного и повторного привыкания. Если это так, то как это происходит? Наконец, каким образом изменение в структуре нервного импульса, вызываемого привыканием, действует на микроструктуру медленного потенциала на следующем синаптическом уровне? Чтобы ответить на некоторые из зтих вопросов, рассмотрим нейронную организацию сетчатки, этого маленького «кусочка мозга», который является гораздо более доступным для изучения, чем центральная нервная система.
АДАПТАЦИЯ
То, что мы воспринимаем из нашего окружения, зависит от возможностей наших рецепторных органов, которые накладывают свои ограничения на восприятие той или другой конфигурации воздействия. Так, оптическая система глаза фокусирует на сетчатку ограниченный диапазон электромагнитных волн; звуковые волны оказывают давление на жидкость в улитке уха; разного рода деформации кожи возбуждают свободные или специализированные нервные окончания в соматической системе и т. д. Эти формы энергетических изменений взаимодействуют «собственной активностью рецепторов и вызывают ее изменения, которые являются достаточно надежными, чтобы организм мог их идентифицировать как результат внешних воздействий.
Воспользуемся еще раз простой моделью протекания речевого акта, описанного в гл. I. Там участвовали две основные переменные: устойчивое состояние и дискретные переменные. Эта модель была применена к функции мозга, сейчас мы используем ее для сенсорного механизма. Заменим состояние постоянного напряжения голосовой связки постоянным распределением различных видов энергии по рецепторным поверхностям, дискретные колебания воздуха — дискретными нейронными разрядами в форме нервных импульсов. Короче говоря, предположим, что то, что происходит в органе чувств, совсем не отличается от того, что происходит в нервной системе, и что нейрофизиология сенсорных процессов может служить миниатюрной моделью ориентировочного рефлекса и процесса привыкания.
Всем известен процесс адаптации: ощущение при погружении в слишком горячую ванну и осознание спустя несколько минут, что для полного удовольствия следовало бы добавить еще теплой воды; исчезновение ощущения давления на кожу, ожидание
в течение нескольких минут после входа в темный зал кинотеатра,, прежде чем мы что-либо увидим и будем способны найти свободное место. Можно привести много примеров сенсорной адаптации. Наиболее яркий из них тоже парадоксален и был получен в экспериментах, при которых проецируемый на сетчатку образ стабилизировался с помощью зеркал и линз (Ditchburn and Ginsborg, 1952; Riggs, Rattliff, Cornsweet and Comsweet, 1953).
Удивительно, что наши глаза находятся в постоянном движении — даже тогда, когда мы фиксируем точку. Эти небольшие, подобные тремору движения глаз можно зарегистрировать. Такие движения у некоторых людей настолько велики, что заметны другим, но — и в этом-то и состоит парадокс — человек с такими усиленными движениями глаз не знает о них до тех пор, пока не обратит на них внимание, когда смотрит на себя в зеркало (что обычно заставляет его обратиться к врачу, который, если он знаком с такой аномалией, успокаивает своего пациента, говоря, что это не опасно). Движения глаз препятствует тому, чтобы каждый из рецепторных элементов в течение какого-то отрезка времени возбуждался бы одним и тем же соотношением света и темноты, конечно, за исключением таких ситуаций, как плотный туман, когда свет теряет форму и функция зрения сводится к различению яркости. Чтобы изучить последствия нарушений таких движений глаз, на склере, белой части глазного яблока, не обладающей чувствительностью, укрепляют зеркало. Изображение проецируется на зеркало, отражается через призму на гладкую поверхность экрана, на которую смотрит наблюдатель. Призма корректирует отклонения рассматриваемого объекта, соответствующие отклонениям глазного яблока. Благодаря этому изображение, проецируемое на поверхность, всегда падает на одно и то же место сетчатки и образ стабилизируется (рис. Ш-3).
Стабилизированное изображение быстро становится незаметным. Зрительный прибор за несколько секунд так адаптируется, что изображение не может оставаться видимым — оно исчезает, адаптация завершена. Сходный опыт можно провести и в кожно-кинестетической системе: положите предмет на руку и некоторое время подержите его. Скоро ощущение наличия объекта исчезает.
Если бы не было такого механизма, организм подвергался бы непрерывной бомбардировке раздражителей разной длительности и интенсивности и это сделало бы его неспособным к тонкому различению. Фактически возможность зрительного различения, несмотря на изменение фонового освещения в диапазоне от 10 миллиардов до единицы, составляет одно из тех противоречий, которые вызывают большой поток исследований. В данном случае исследования адаптации сетчатки привели к открытию соответствующих нейронных механизмов. Это противоречие
получило объяснение в концепции, согласно которой адаптация сетчатки целиком объясняется выцветанием и регенерацией фоточувствительного пигмента, содержащегося в рецепторах сетчатки (Hecht, 1934). Однако в последнее время стали накапливаться данные, говорящие о том, что для объяснения процесса адаптации необходимы и нефотохимические факторы я что эти факторы являются основой для осуществления фотохимических процессов (см. Rushton, в обзоре Dowling, 1967).
Главным инструментом в этих и< следованиях служит большой электрод, который помещают па внешней стороне глаза.
Рис. Ш-3. Благодаря тому, что стимул, прежде чем воздействовать на сетчатку, первоначально отражается от зеркала, укрепленного на склере и перемещающегося вместе с движениями глаз, последние могут быть компенсированы с такой точностью, что изображение раздражителя на сетчатке становится стабилизированным (см. Riggs et al, 1953).
Этот электрод записывает изменения потенциалов, генерируемых всей сетчаткой, в виде электроретинограммы. При соответствующих методах анализа получают такие же четкие ответы, как и тогда, когда используют микроэлектродную технику.
Коротко говоря, сетчатка млекопитающего состоит из нескольких слоев: рецепторный слой образован клетками, имеющими •форму палочек и (или) колбочек, в которых имеются фоточувствительные химические пигменты; реагирующий первичный слой состоит из клеток, в которых процессы распространяются торизонтально и связывают друг с другом множество соседних клеток сетчатки; биполярный слой состоит из клеток, связывающих один или несколько рецепторов с ганглиозными клетками, являющимися началом выхода импульсов из сетчатки к мозгу <рис. Ш-4).
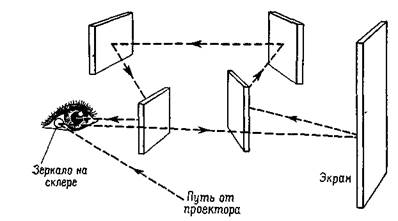

Рис. Ш-4. Схема строения сетчатки приматов, показывающая синапсы между различными типами клеток, наблюдаемые с помощью электронного микроскопа на серийных срезах. Обратите внимание на контакты между рецепторами, на широко распространенные связи горизонтальных и ама-криновых клеток и вертикальное расположение биполяров. Вздутые центральные окончания биполярных клеток формируют так называемые диад-ные синапсы (в рамке) с гаяглиозной клеткой и одновременно с амакри-новой клеткой. П — палочка; К — колбочка; кб — карликовый биполяр; по — палочковый биполяр; плб — плоский €иполяр: г — горизонтальная клетка; а — амакриновая клетка; кг — карликовая ганглиозная клетка; д — диадный синапс; дг — диффузная ганглиозная клетка; н — ножка. В прямоугольной рамке вверху справа показаны детали типичною диадного синапса (Horridge, 1968; Dowling and Boycott, 1966).
Электроретинограмма включает в себя два компонента: малую а- и большую e-волну; а-волна, по-видимому, генерируется более периферической частью сетчатки, чем e-волна. Это обнаруживается при наложении на зрительный нерв зажима. В результате этой операции нарушается кровообращение в сетчатке, за исключением тех сосудов, которые питают рецепторы. В таком препарате остаются только а-волны.
Сходную процедуру можно применить и для разрушения ганглиозных клеток, образующих самый глубокий слой сетчатки; рассечение зрительного нерва вблизи его начала вызовет дегенерацию большинства тел порождающих импульсы ганглиозных клеток. Электроретинограмма в результате этой процедуры, по-видимому, не нарушается; фактически адаптация в таких препаратах остается нормальной. Таким образом, этот самый глубокий слой сетчатки не может быть источником ни а- ни, e-волн. Это -значит, что генераторами e-волны являются средние слои сетчатки.
Процесс адаптации для а- и e-волн различен. Адаптация а- волны не отражает хода психофизической адаптации, и поэтому мы не будем рассматривать ее здесь подробно. Напротив, адаптация e-волны происходит параллельно психофизической адаптации и ее механизм является для нас центральным.
Ряд остроумных экспериментов принес подтверждение концепции, согласно которой адаптация является функцией не отдельных клеток сетчатки, а целой группы нейронов, составляющих нейронную сеть сетчатки. Типичный эксперимент показывает, что чувствительность к свету остается одной и той же, когда на сетчатку проецируется рисунок в виде чередования темных и светлых полос или когда она освещается светом той же средней интенсивности, но равномерно распределяемым по ловерхности. Этот тип исследований находит логическое завершение в эксперименте, в котором было показано, что освещение одной части зрительного рецептивного поля одиночной ганглиозной клетки (определяемого с помощью микроэлектрода) вызывает процесс адаптации в другой его части.
Множество данных говорит о том, что биполярные клетки и клетки, осуществляющие взаимодействие в среднем слое сетчатки, являются генераторами e-волны и местом психофизической адаптации. Некоторые исследователи (например, Fuortes, Hodgkin, 1964; Rushton, 1963; Dowling, 1967) считают, что нейронная адаптация возникает в результате действия механизма обратной связи, когда сигнал с одной ступени посылается вазад на предыдущую ступень и таким образом снижает ее •чувствительность. Недавно Дж. Даулинг и Б. Бойкотт (1965)' показали с помощью электронного микроскопа, что контакты биполярной, амакриновой и ганглиозной клеток могут функционировать точно таким же способом. Были открыты реципрокные
синапсы, они делают возможным движение возбуждения назад", от амакриновой клетки к биполярной, от которой амакриновая клетка ранее получила возбуждение. Подразумевается, что эта обратная связь является отрицательной, что создает систему сер-

Р и с. Ш-5. Зарисовка нейрона в зрительной системе мозга (латеральное коленчатое тело), иллюстрирующая синаптические контакты. Обратите внимание, что, как и в сетчатке, пузырьки, содержащие химический возбудитель, находятся иногда на дальней стороне, иногда — на ближней стороне синаптической щели, показывая, что нейрон посылает сигналы и воспринимает их (обозначено направлением стрелок). Эта двойная направленность связей (диадные синапсы) делает возможной обратную связь-
(Ralston, 1971).
вомеханизма, но это утверждение нуждается в более прямом доказательстве (рис. Ш-5).
УСИЛЕНИЕ КОНТРАСТА
Затухание нейронных ответов само по себе не может объяснить, каким образом формируется «нейронная модель» памяти,, с которой сравниваются последующие воздействия. Как мы уже говорили, полученные при изучении поведения данные свидетельствуют о том, что даже при малейшем изменении сложного
стимула, к которому организм обнаружил привыкание, возникает растормаживание. Необходима определенная преобразующая организация нейронных явлений. Как же осуществляется такая организация?
Чтобы ответить на этот вопрос, рассмотрим одно из тех противоречий, которыми так богата наука о мозге. В данном случае это противоречие было замечено венским физиком Эрнстом Махом. Мах отметил, что, когда человеку предъявляется более или менее неравномерно освещенная плоскость, воспринимаемые различия в яркости увеличиваются. В то время как физическое изменение, замеренное с помощью прибора, можно описать как

восприятие изменения выглядит более похожим на (см. рис. III-6).

Другими словами, кажется, что плоскость имеет в местах изменения освещения темные и светлые «полосы», известные под именем «полос Маха». Можно предположить, что психофизическое несоответствие объясняется тем, что зрительный аппарат (вероятно, сетчатка) функционирует, чтобы дифференцировать, в математическом смысле, интенсивность света в соответствии с тем, как свет распределяется по ее поверхности. Согласно этому объяснению, восприятие зрительного контраста обусловлено, до-видимому, нейронным механизмом, выполняющим функцию дифференцирования. Математическая модель была улучшена и модернизирована при более детальном анализе психофизических порогов различения. Этими исследованиями были подтверждены также правомерность подхода Маха, а также некоторые элементы его решения этой проблемы. Вместе с тем они представили и нейрофизиологические данные о механизме, ответственном за зрительный контраст (а на основе экстраполяции — и за контур).
Путем вживления микроэлектрода в нервное волокно, берущее начало от ганглиозной клетки, экспериментатор может создать карту зрительного поля, — вернее, находящейся перед глазами области, — в которой преходящее появление света будет вызывать изменение в частоте импульсных разрядов этой ганглиозной клетки. Такая карта известна как зрительное рецептивное поле (см. рис. 1П-7). Выбирая различные клетки, можно получить ряд карт: большинство карт более или менее круглые-
по форме, но некоторые из них звездообразные, или линейные, или имеют длинный край, отделяющий часть рецептивного поля, в котором свет вызывает этот эффект, от части, где эффект отсутствует. Вообще можно выделить два класса зрительных рецептивных полей: рецептивные поля, у которых свет в начале тормозит разряды ганглиозных клеток, и рецептивные поля, реагирующие на свет усилением спайковых разрядов нейрона. Далее, каждое из первично реагирующих полей частично или
полностью окружено другой областью, освещение которой вызывает у клетки ответ с противоположным знаком. Таким образом, большинство ганглиозных клеток можно разделить на единицы с оп- эффектом или о//-эффектом в центре; ore-центральные единицы характеризуются, как правило, наличием тормозного окружения; о//-центральные единицы часто окружены зоной, реагирующей возрастанием частоты разрядов. Эти карты зрительных рецептивных полей ганглиозных клеток указывают на функциональную организацию сетчатки. Очевидно, такие записи являются не просто отражением активности отдельных рецепторов. Ганглиозные клетки — это нейроны третьего порядка, которые активируются любым большим числом рецепторов, с которыми они анатомически связаны. Рецептивные поля в большой степени соответствуют размерам фоторецепторов, и соседние поля значительно перекрывают друг друга. Таким образом, любая отдельная область рецеп-торной мозаики не связана исключительно с какой-нибудь одной особой ганглиозной клеткой (Ratliff, 1965, р. 173—174).
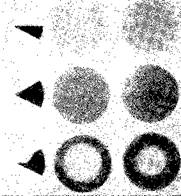
Наличие в рецептивном поле зоны с эффектом, противоположным по знаку центральному эффекту, означает, что антагонистические возбуждающие и тормозные влияния организованы таким образом, что стимуляция рецепторов одного участка снижает эффект возбуждения, получаемого ганглиозной клеткой от соседних стимулированных клеток. Другими словами, возбуждение какого-то участка сетчатки продуцирует торможение вокруг этого участка. Этот процесс «периферического», или «латерального», торможения непосредственно наблюдается в глазе подковообразного краба Limulus (рис. III-8). У этого животного элементы
Рис. Ш-6. Вращение диска с изображенными на них картами (левая колонка) формирует стимулы, физическая и субъективная характеристика которых показана в средней и правой колонках (см. «Psychology Today», 1970).
сетчатки пространственно отделены друг от друга — каждый элемент образует свою собственную, заключенную в капсулу единицу, названную омматидием. Поэтому при микроэлектродном исследовании волокон, выходящих из сетчатки, можно отдельно' осветить каждый омматидий. Освещение одного из таких оммати-диев будет вызывать возбуждение, если запись осуществляется

Рис. Ш-7. Точечная карта сетчатки, в пределах которой световое пятно вызывает ответ определенного нейрона латерального коленчатого тела в мозгу обезьяны (Spinelli and Pribram, 1967).
от волокон, чье рецептивное поле включает этот омматидий. Когда свет перемещается на соседний омматидий, микроэлектрод регистрирует торможение. Полученные таким образом карты сходны с картами ганглиозных клеток млекопитающих, построенных в. результате сканирования рецепторов в пределах их рецептивных полей методом короткой вспышки света.
Этот процесс «периферического», или «латерального», торможения не ограничивается зрительной системой. Сходную функциональную организацию имеют кохлеарный механизм слуха и рецепторы кожи (Bekesy, 1967). В центральной нервной системе аналогичным образом реагируют клетки коры головного мозга и коры мозжечка.
Короче говоря, латеральное торможение является одной нехарактерных особенностей нейронных сетей — особенно тех, кото-

0 12 3
Время (сек)
Тис. III-8. Одновременное кратковременное развитие возбуждения и торможения у двух соседних рецепторных единиц в латеральном глазе Limu-lus'a. Одна рецепторная единица (черные кружки) постоянно освещалась в течение всего периода, показанного на графике. Освещение другой единицы (белые кружки) также оставалось на одном постоянном уровне, кроме отрезка времени от 0 до 2 сек, когда ее освещение резко возрастало и оставалось на новом постоянном уровне. Заметный кратковременный рост возбуждения у одной рецепторной единицы сопровождается большими кратковременными тормозными эффектами у соседней, постоянно освещенной единицы. Резкое падение частоты вызывается тормозным эффектом, возникающим в результате большого кратковременного возбуждения. Во время постоянного освещения тормозный эффект еще присутствует, но менее заметен. Наконец, затухание частоты ответа у элемента, у которого уровень возбуждения был снижен, сопровождается заметным высвобождением от торможения (Ratliff, 1965).
рые организованы в плоские слои. В этих слоях располагается несколько уровней обработки информации о сигналу.
Какие же свойства этих нейронных сетей объясняют латеральное торможение? Было дано несколько объяснений латеральному торможению, и они не являются взаимно исключающими; точное ^математическое) описание и оценку каждого из них читатель
может найти в обзоре Рэтлиффа (1965, гл. 3). Нуждается в •объяснении прежде всего возникновение латерального торможения, его явная зависимость от расстояния до центра возбуждения, очевидность взаимодействия между возбуждением и торможением. Наиболее правдоподобное объяснение состоит в том, что разветвления каждого рецептора образуют тормозные связи с соседними рецепторами или, что более вероятно, они возбуждают
Входы (раздражители)
Выходы (импульсы)
Рис. Ш-9. Схема рецепторного слоя и слоя взаимодействия (горизонтальные клетки) в сетчатке. Вертикальные стрелки указывают направление передачи сигнала, горизонтальные — тормозные взаимодействия (Ratliff, 1965).
тормозные нейроны — например, амакриновые и горизонтальные клетки сетчатки, обширные разветвления дендритов которых и отсутствие у них аксона делают их идеальными для выполнения этих функций (рис. Ш-9). Обнаружено, что такие не имеющие аксона дендритные сети в сетчатке (Svaetichin, 1967) и в каком-нибудь другом месте нервной системы (например, в мозжечке) выполняют точно такую же функцию. Действительно, недавние эксперименты (Werblin and Dowling, 1969) с использованием внутриклеточной регистрации показали, что горизонтальные клетки функционируют исключительно посредством гиперполяризации, то есть путем генерации тормозных медленных потенциалов. (Фактически от элементов сетчатки, расположенных на периферии слоя ганглиозных клеток, можно зарегистрировать только медленные потенциалы, а не нервные импульсы.) А возникновение торможения за счет дендритных сплетений, в которых медленные потенциалы генерируются в ответ на

воздействия входных волокон, выявляя рисунок продуцируемых волновых форм, по существу, ведет к созданию эффектов интерференции, которые в их простейшей форме и образуют периферическое торможение.
В заключение следует отметить, что сенсорный механизм почти постоянно изменяет свою реакцию на входное воздействие, используя для этого два процесса: усиление контраста и адаптацию. Анатомические связи механизма делают вероятным реципрокное функционирование этих процессов; чем больше ослабевают ответы нейрона в ходе адаптации, тем меньшее тормозное
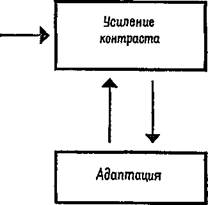
Рис. Ш-10. Реципрокные отношения усиления контраста и адаптации.
влияние оказывает он на соседние нейроны, вызывающие усиление контраста (рис. Ш-10).
Таким образом, прогрессивно растущее торможение, затухание возбуждения в нервной системе препятствует развитию длительного разряда, делает возможным последовательное сравнение входных структур возбуждения и до некоторой степени усиливает временный контраст путем простого механизма вычитания. Далее, тормозные взаимодействия между соседними нейронами препятствуют распространению, или иррадиации, возбуждения по рецепторным сетям — тормозные взаимодействия усиливают четкость, очерченность пространственных узоров. Таким образом, проведение информации о контрасте на системах входа осуществляется более надежно, чем если бы для этой цели использовались предварительно не обработанные данные об энергетических изменениях, как таковых, воздействующих на входные пути нервной системы.
Эти явления адаптации и усиления контраста свойственны не только рецепторам. Взаимодействия, ведущие к ослаблению и
торможению ответа, имеют место во всех системах входных путей, и особенно в коре головного мозга (Brooks and Asanuma, 1965). Это подтверждает ранее высказанное предположение, что наблюдаемые в рецепторах процессы адаптации и контрастирования могут служить миниатюрными моделями психологических процессов ориентировочного рефлекса и привыкания.
В следующей главе мы покажем, как эти элементарные процессы вместе с другими, имеющими еще более короткую продолжительность, приобретают такую важную роль и пронизывает все виды деятельности организма.
РЕЗЮМЕ
Процессы мозга, характеризующиеся средней продолжительностью, являются в значительной мере результатом развития у нейронов процессов затухания и торможения. При однообразной стимуляции у многих групп нейронов наблюдается ослабление активности (адаптация и привыкание) и они, таким образом, становятся чувствительными к новизне (ориентировочная реакция). Во многих частях нервной системы локальное возбуждение нейронов тормозит активность окружающих нейронов и тем самым усиливает контраст между возбужденной и невозбужденной тканями. Процесс затухания у нейронов, видимо, развивается в системе генерации потенциала, который посредством деполяризации порождает нервные импульсы; тормозные же взаимодействия зависят от гиперполяризаций, возникающих в соединительной сети.
Глава IV
КОДЫ И ИХ ПРЕОБРАЗОВАНИЯ
ЧТО ТАКОЕ КОД?
Как можно охарактеризовать формы тех временных нейро-электрических конфигураций (и отношений между ними), которые делают возможным существование самих механизмов кратковременной памяти, механизмов, на какие мы опираемся, когда имеем дело, например, с конструкцией немецкого предложения с глаголом на конце? Изучение условий, влияющих на человеческую память, продемонстрировало несколько пренебрежительное отношение к проблеме структур; если что-либо запоминается,, то это объяснялось в значительной мере формой материала и контекстом, в котором он предъявлялся. Мы же обращаемся к проблеме замещения одной конфигурации другой, которое возникает в результате определенных операций в нервной системе. На техническом языке — это проблема преобразований или функций передачи, что делает возможным кодирование и перекодирование.
Операции кодирования постоянно имеют место в нервной системе. Физическая энергия воспринимается рецепторами и трансформируется в нервные импульсы. Эти импульсы в свою очередь достигают синаптических сетей, где дискретные сигналы кодируются в микроструктуры медленных потенциалов. Для того чтобы кодирование было эффективным и могло быть использовано мозгом, необходимо наличие декодирующей операции, благодаря которой было бы возможно восстановление нервных импульсов и сохранение, таким образом, предварительно закодированной информации.
Для описания операций кодирования, происходящих в нервной системе, можно использовать два класса функций передачи.
Одни преобразования допускают достаточно простой расчет соответствия между кодами. Такой расчет делает возможным декодирование закодированной формы, и наоборот. Благодаря
однозначному соответствию между цифрами кода между ними устанавливается нечто вроде обратимости, вторичный функциональный изоморфизм.
Второй класс функций передачи целиком неизоморфный и условный. Преобразования в этом классе необратимы, пока нет ключа для дешифровки такого кода.
Обратимые трансформации возникают в физико-оптических системах, в звуковых преобразователях и, как мы покажем, в закодированных нейронных конфигурациях микроструктуры медленных потенциалов мозга. Необратимые преобразования имеют место в обычных языках, состоящих из условных алфавитных комбинаций, в азбуке Морзе, в телефонной и телевизионной передаче сигналов и т. п. В нервной системе, как будет подробно рассмотрено в последующих главах, имеют место необратимые преобразования, если происходит процесс абстрагирования, например такой, как детекция признака.
Для чего мозгу нужно так много замещающих схем, так много операций кодирования и перекодирования? Ведь любое преобразование грозит утратой точности. Почему же это свойство присуще всей нервной системе? Какую пользу дает перекодирование? Должно быть, ответ на этот вопрос состоит в том, что перекодирование повышает эффективность нервной системы.
Это перекодирование отнюдь не такая тривиальная операция, как та, с которой я столкнулся при использовании компьютера общего типа. Последним можно управлять только с помощью языка, который он понимает: пространственной или временной последовательностью «on» и «off», «да» и «нет», «вверх» и «вниз», системой имеющихся у него переключателей. Если имеется 12 таких переключателей, то мы должны запомнить размещение каждого из них для осуществления операции, которую мы хотим получить от компьютера. Следовательно, передача сообщения принимает вид серий
ABB AAA BAA BBB ААВ ABA AAB ABB ABA BBB ABA ABA и т. д.
Таким образом, стоящая перед оператором компьютера задача является в значительной мере задачей на запоминание длинного перечня позиций включения и выключения, рядов «двоичных» цифр.
• 011 000 100 111 001 010 001 011 010 111 010 010
Программисты должны были быстро овладеть этим сложным способом управления их инструментом: они подразделяли 12 переключателей на наборы по три и обозначали включение (В) целым числом, геометрически увеличивающимся справа налево..
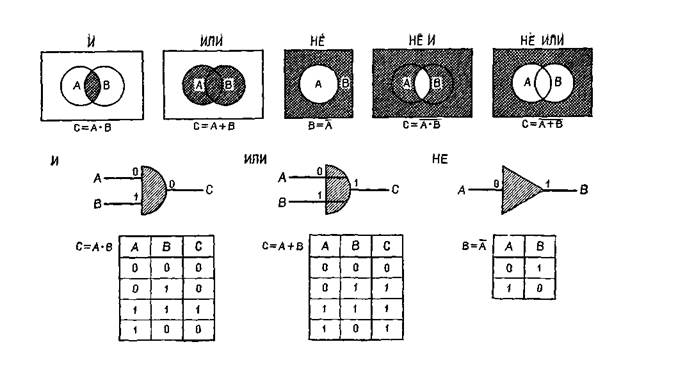
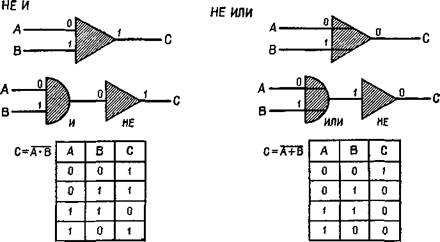
Рис. IV-1. Представление двоичных логических элементов: «и», «или», «не и», «не или». Входные сигналы 0 или 1 поступают в схему из двоичных элементов слева, где логически комбинируются для создания выходного сигнала справа. «Таблица соответствий» под каждой схемой перечисляет все возможные логические операции, которые могут еще быть выполнены. Элементы «не и» и «не или» образуются комбинациями элемента «не» с «и» и «или» (Evans,
Computer Logic and Memory, 1966).
ДЕСЯТИЧНАЯ ДВОИЧНАЯ
| Аз | А2 | А< | Ао | |
| О | ||||
| о 2 | ||||
| f | ||||

Рис. IV-2. Превращение двоичной системы в десятичную с помощью схемы, состоящей из 4 элементов «не» и «и». Таблица соответствий справа содержит двоичный эквивалент для десятичных цифр (от 0 до 9). Схема демонстрирует принцип декодирования двоичных чисел на примере декодирования числа 2. Сигнал на каждом из пронумерованных выходов будет равен 0, пока на все входы подается 1. В этом примере единица подается на вход А! третьего справа логического элемента «и», обозначенного цифрой 2. Таким образом двоичное число 0010 декодируется в число 2 десятичной системы (см. Evans, 1966).

Рис. IV-З. Схема сетчатки приматов, показывающая типы нейронов и их синантические связи Метод Гольдзди. (Polyak, 1941).
Таким образом, в каждой триаде включение указывает на 4—2—1. Когда одновременно включаются два или три переключателя, то они представляют сумму целых чисел. Так,
0 означает AAA
1 означает ААВ
2 означает ABA
3 означает ABB
4 означает ВАА
5 означает ВАВ
6 означает ВВА
7 означает ВВВ
и любую последовательность из 12 включений и выключений можно описать и запомнить в виде четырех цифр, например представленные ранее последовательности превращаются в 3047,
1213, 2722. Это преобразование, названное «восьмиричным» кодированием двоичной системы, удивительно экономно. Каким же образом стало возможно столь замечательное достижение? (Этот -совершенный способ нелегко приобретается нервной системой; благодаря повторению процесса обеспечивается классификация по иерархическим схемам, а классификация — наиболее фундаментальная логическая процедура.) Чтобы получить ответ, сформулируем проблему более конкретно: каким образом может происходить преобразование одной структуры, сложность которой яредставлена системой связей между простыми элементами, в другую структуру, сложность которой определяется самими элементами и их уникальным назначением? Такое преобразование может быть осуществлено с помощью простой системы конвергентных единиц, названных инженерами функциями «и», так как их реакция возникает в ответ на одновременный приход •сигналов (см. рис. IV-1 и IV-2). Такую систему пересекают пути, выполняющие функцию параллельного торможения — «не или», — которая реализуется через комбинацию функций «или» (у которых выходной сигнал определяется воздействием через любой из двух входов) и отрицательной функцией «не», тормозящей выходной сигнал элемента, когда каким-либо образом активируется •«го вход. Эта система так напоминает структуру сетчатки — наше -окно в мозг, что испытываешь искушение назвать функцией «не или» гиперполяризацию горизонтального слоя и функцией «и» — разряд ганглиозной клетки. Конечно, сетчатка построена не вполне так, как показано на этой схеме, однако обе формы •организации поразительно соответствуют друг другу с точки прения рассматриваемого процесса (ср. рис. IV-2 и IV-3).
Таким образом, перекодирование оказывается чрезвычайно эффективной частью процесса памяти, для осуществления которого конструкция нервной системы, по-видимому, великолепно.приспособлена. Формы перекодирования, которые возможны в нервной системе, фактически безграничны. Тем не менее можно выделить несколько классов кодов.
ТИПЫ НЕРВНЫХ КОДОВ
Мы уже разделили коды на два класса: коды, предполагающие дискретные импульсы нервного разряда, и коды, предполагающие микроструктуру устойчивого состояния, создаваемую нервными соединениями.
Проблема импульсного кодирования не сводится к вопросу оприсутствии или отсутствии сигнала; импульсный код может использовать множество различных параметров, которые характеризуют последовательности залпов.

Рис. IV-4. Кодирование интенсивности и места раздражения, осуществляемое параллельными волокнами. Пространственное представительство показано в соответствии с тем, что было обнаружено* в тактильных афферентах задней лапы кошки, когда последняя возбуждалась механическими залпами, приложенными к одной точке; диаграмма основывается на количественных экспериментальных данных. Если число равноотстоящих рецепторов с 3 уровнями возбудимости соответствует схеме е, то раздражитель интенсивностью в два порога вызывает некоторое число импульсов в стороне от раздражаемой точки и через какое-то время после раздражения а. Пунктирная линия обозначает латентный период, обусловленный временем распространения механической волны по лапе. Более сильные раздражители вызывают ответы, показанные на рис. б-г. Общее число импульсов в ответ на каждую интенсивность раздражителя представлено в д (Gray and Lai, 1965).
Д. Перкел и Т. Буллок (Perkel and Bullock, 1968), изучал сигналы, посылаемые электрическими органами некоторых рыб, установили следующие типы кодирования. При той или: иной комбинации условий изменяется длительность латентного
периода, продолжительность спайковых взрывов, общая вероятность разрядов и вариация этой вероятности, увеличивается или уменьшается частота разрядов или скорость ее изменения. Такими условиями являются искажения электрического поля, создаваемые теми же самыми сигналами, посылаемыми электрическими органами, — искажения, регистрируемые системой боковой линии рыбы, этот процесс сходен с радарным.
Кроме этих организованных во времени кодов, имеет место и пространственное кодирование, когда множество параллельных линий — нервных волокон — продуцируют то, что Перкел и Буллой назвали обработкой «по ансамблю», которая зависит от различий в распределении последовательностей импульсов вдоль линий (см. рис. IV-4).
Пространственное кодирование и особенно кодирование, опирающееся на различия в синхронности разрядов у соседних групп нервов, тесно связано с целой группой процессов неимпульсного кодирования — микроструктурами медленных потенциалов, обеспечивающих устойчивое состояние, которые вносят существенный вклад в активность серого вещества мозга млекопитающих. Как говорят Перкел и Буллок, должно существовать какое-то считывающее устройство, какой-то механизм в центральной нервной системе, который считывает сообщения, закодированные в импульсной форме. В этих главах утверждается, что такое считывание возникает на местах соединений нейронов в процессе образования микроструктур медленных потенциалов, мгновенных состояний, возникающих в результате взаимодействия между совокупностями сигналов, действующих по соседству и кодируемых последовательностью импульсов.
СПОНТАННАЯ АКТИВНОСТЬ НЕРВНОЙ ТКАНИ
Чтобы быть эффективными, большинство кодов нуждается в некотором стабильном уровне Работа Бернса и других, вне веяного сомнения, установила, что активность центральной нервной системы обладает такой стабильностью Нервная ткань спонтанно генерирует электрические потенциалы Мозг, как и сердце, непрерывно пульсирует И, так же как в сердце, такую пульсацию вызывают медленные потенциалы, а возникновение этих последних зависит от определенных констант химического окружения, в котором находится пульсирующая 1кань (рис IV-5).
Серии тщательно проведенных исследований в лаборатории Бернса (1958) дали исчерпывающий ответ на вопрос, который долгое время оставался фантастическим: может ли мозг сохранять активность даже в том случае, если он полностью изолирован (нейронально) от другой нервной ткани? Результаты этих экспериментов, как это часто бывает, не подтвердили полностью
Рис IV-5. Церебральная симфония (Verzeano et al, 1970).

ни представления о том, что активность головного мозга «спонтанна», ни представления о мозге как находящейся в покое tabula rasa, на которую записывается сенсорный опыт. Берне обнаружил, что даже у неанестезированного животного изолированная полоска коры остается неактивной до тех пор, пока к ней хотя бы на короткое время не будет приложено электрическое раздражение; другие же данные (Echlin et al., 1952; Gerard and Joung, 1937; Henry and Scoville, 1952; Ingvar, 1955; Libet and Gerard, 1939) указывают на то, что спонтанная активность существует и в таких препаратах. В любом случае, даже если принять осторожный вывод, сделанный Бёрнсом, несколько сильных электрических раздражителей, приложенных к поверхности коры, вызывают серию разрядов нейронной активности, которая обычно продолжается в течение многих минут (или даже часов) после прекращения стимуляции.
Периодические волны возбуждения могут быть получены и в диффузно организованной нервной ткани при ее электрическом раздражении. Они сходны с волнами возбуждения, возникающими в неанестезированной коре головного мозга в ответ на воздействие нескольких редких стимулов. Эффекты, длящиеся многие часы, наблюдались после короткой стимуляции интактной актинии (Batham and Pantin, 1950). Недавно был описан люминесцентный ответ у морских «анютиных глазок» (вид цветного коралла): после серий раздражений эти колонии начали люминесцировать спонтанно, а не только в ответ на стимуляцию. Для объяснения этого явления следует обратиться к механизму медленных изменений состояния нервной ткани (элементарной форме памяти, связанной с медленными потенциалами?) Эти изменения обусловлены влиянием окружающей среды и зависят, разумеется, от предшествующей активности организма. Но они также имеют свои внутренние закономерности и свой собственный ритм активности, который вызывает повторные изменения состояний нервной ткани, что делает их в каждый момент времени лишь частично зависимыми от влияний окружающей среды.
Короче говоря, принято считать, что группы нейронов того типа, которые найдены в коре головного мозга, в отсутствие непрерывного сенсорного воздействия находятся в состоянии покоя. Однако эти группы нейронов могут легко приходить в состояние- возбуждения и обнаруживать длительную активность. Значит, можно считать, что во время «покоя» они находятся в состоянии ниже порога непрерывного самовозбуждения. У интактного млекопитающего есть механизм, который поддерживает возбуждение центральной нервной системы выше этого уровня покоя. Таким механизмом является спонтанный разряд рецепторов.
Р. Гранит (1955) подробно рассказал о том, каким образом им «завладела мысль, что спонтанная активность является со-
ставной частью работы сенсорных систем». Он проследил историю этого вопроса от ранних наблюдений лорда Э. Эдриана и И. Зоттермана (1926), Э. Эдриана и Б. Мэттьюза (1927а, б), выполненных на мышцах и препаратах зрительного нерва, до его собственных разносторонних экспериментальных исследований. Более того, его данные подтверждают предположение, что эта «спонтанная» активность органов чувств делает их одним из наиболее важных «энергизаторов», или активизаторов, мозга. Сейчас мы можем добавить к этому, что, вероятно, эта спонтанная активность является той основой, тем уровнем, на котором и по отношению к которому осуществляется нейронное кодирование. Берне также представил данные, подтверждающие такое предположение (1968). С помощью микроэлектродов он обнаружил, что примерно 7з большого числа обследованных им клеток мозга в течение всего времени, пока он вел от них запись, показала устойчивость средней частоты своих разрядов. Эти нейроны реагировали на стимуляцию либо возрастанием частоты разрядов, либо их торможением. Всякий раз вслед за этим наступал период, во время которого активность нейрона менялась реципрокно реакции. В результате происходила компенсация изменений средней частоты разрядов нейрона, вызванных стимуляцией. Таким образом, эти клетки создают мощную стабильную базу, от которой зависит основная характеристика кодирования и перекодирования: пространственные структуры возбуждения могут возникать за счет возрастания спонтанной активности в одном месте и одновременного ее торможения в другом.
Date: 2016-05-25; view: 488; Нарушение авторских прав