Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Теории механизма мышечного сокращения
|
|
До получения данных о тонкой структуре мышц процессы мышечного сокращения пытались объяснить деформацией изолированных молекулярных цепей белков, т. е. удлинением или укорочением отдельных белковых молекул или агрегатов молекул. Часто данные о деформации различных полимеров переносили на мышечное сокращение, без учета структуры мышечных волокон.
Существует множество гипотез, пытающихся объяснить мышечное сокращение на основе свойств индивидуальных молекулярных цепей сократительных белков. Все эти гипотезы исходят из представления, что в основе сокращения мышцы лежат процессы конформационных изменений структуры белковых цепей. Так, Мейер еще в 1929 г. выдвинул гипотезу, согласно которой мышечное сокращение обусловлено деформацией пептидных цепей вследствие изменения электростатического взаимодействия ионогенных групп СООН и NH2 при изменении рН.
В настоящее время считают, что изменение рН при возбуждении миофибрилл недостаточно, чтобы вызвать конформационные переходы белков, по может быть достаточно для освобождения ионов кальция, которые уже могут вызвать деформацию белковой цепи.
Однако эти гипотезы не смогли объяснить реальную картину сложного строения мышечного волокна на молекулярном уровне, полученную в последнее время. Возможно, что при медленном сокращении гладких мышц происходит фактическая деформация (активное сокращение отдельных протофибрилл) белковых цепей, как считает Г.М.Франк, однако для сокращения скелетных мышц гораздо более обоснованными являются представления, исходящие из гипотезы скольжения нитей.
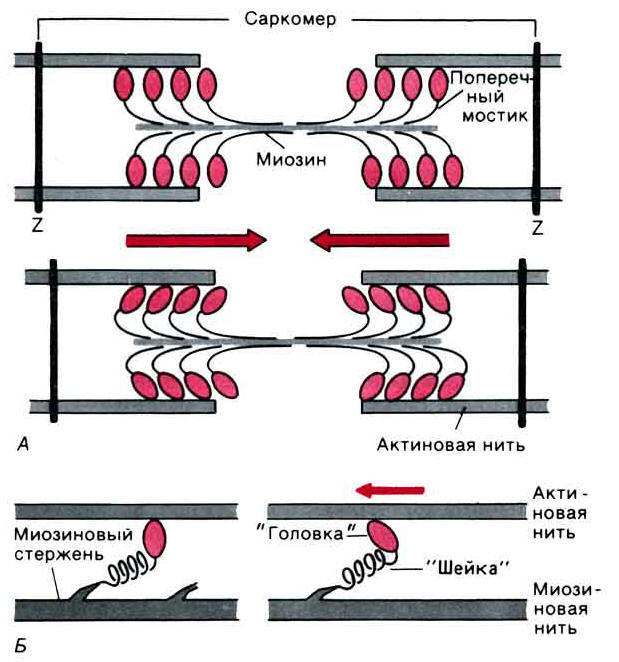
Г.Хаксли и Хэнсон выдвинули гипотезу скольжения нитей. Ими было отмечено, что в широком интервале деформаций как при сокращении, так и при растяжении миофибрилл ширина А-диска остается постоянной. Напротив, при изменении длины саркомера изменяется ширина I-диска. Светлая Н-зона в А-диске также изменяется, но замечательно, что до тех пор, пока она существует, расстояние от конца одной Н-зоны через Z-мембрану до начала следующей Н-зоны (а это расстояние равно длине тонких нитей в миофибрилле) также остается постоянным. Если вспомнить, что А-диски образованы нитями миозина, а тонкие нити состоят их актина, то можно заключить, что в большой области деформаций мышцы длина миозиновых и актиновых нитей остается постоянной. Это можно объяснить только тем, что при сокращении мышцы нити просто скользят друг относительно друга без изменения своей длины.
В дальнейшем данные электронного микроскопирования были подтверждены результатами рентгеноструктурного анализа. Основные рефлексы рентгенограммы не изменяются при сокращении мышц. Это указывает на то, что длина нитей при сокращении не меняется. Приведенные данные очень важны, так как в отличие от электронно-микроскопических исследований, проводимых на фиксированных препаратах мышц, рентгенографические исследования проводились и на живых сокращающихся мышцах, и на нефиксированных ее препаратах.
Перемещение тонких нитей относительно толстых происходит, при помощи мостиков, соединяющих миозиновые нити с актиновыми. Так как изменений в длине толстых и топких нитей во время сокращения не происходит, то из модели скольжения нитей вытекает, что конформационные изменения, порождающие движение, должны происходить в указанных мостиках, связывающих толстые и тонкие нити. Весь процесс сокращения имеет циклический характер. Миозиновые мостики прикрепляются к активным участкам актиновых нитей и под действием энергии гидролиза АТФ укорачиваются или изменяют угол наклона к миозиновым нитям, что приводит к определенному перемещению нитей друг относительно друга. Затем происходит отсоединение мостиков в данных участках актиновых нитей и присоединение их в новых участках. Этот циклический процесс повторяется многократно, в результате чего происходит непрерывное перемещение нитей друг относительно друга. Рентгенографические исследования подтвердили предположение о движении мостиков. По мнению Г.Хаксли, расщепление одной молекулы АТФ приводит к одному замыканию и размыканию мостиков и к перемещению нитей на один элементарный участок.
Величина напряжения, развиваемого мышцей, определяется количеством замыкаемых (функционирующих) мостиков. Если мышца преодолевает при сокращении внешнюю силу, то замыкается такое количество мостиков, которое необходимо для уравновешивания этой силы. Максимальная сила, развиваемая мышцей, определяется количеством мостиков, которые могут замыкаться в данных условиях. Исходя из этих представлений, нетрудно объяснить обратную зависимость напряжения, развиваемого мышцей при сокращении, от скорости сокращения. Для того чтобы мостики замкнулись, необходимо какое-то время. При увеличении скорости скольжения нитей количество замыкаемых мостиков уменьшается, что обусловливает уменьшение напряжения, развиваемого мышцей.
Однако, несмотря на большие успехи в изучении механизма мышечного сокращения, все еще окончательно не установлен механизм работы мостиков, в результате которой энергия гидролиза АТФ превращается в механическую работу.
В настоящее время имеется ряд гипотез, пытающихся объяснить конкретный механизм взаимодействия актиновых и миозиновых нитей.
Наиболее глубоко разработанной и обоснованной является гипотеза Дэвиса. Согласно этой гипотезе, мостик между миозиновой и актиновой нитями образован полипептидными цепочками конца миозиновой молекулы, скрученными в спираль. В покое мостик вытянут-спираль находится в растянутом состоянии. Это обусловлена электростатическим отталкиванием двух отрицательных зарядов. Один из них находится в фиксированном состоянии у основания мостика, которое обладает АТФ-азной активностью. Другой отрицательный заряд локализован па конце мостика, с которым связана молекула АТФ.
Модель Дэвиса получила ряд дополнений и подверглась модификациям. Бендолл (1970) предполагает, что присоединение ионов кальция в области мостиков приводит к изменению электрического взаимодействия. Нейтрализация отрицательных зарядов и присоединение актина к миозину обусловливают превращение спирали полипептидной цепочки (мостика) молекулы миозина в более беспорядочную, сильно свернутую конформацию но типу перехода «спираль - клубок».
Такой переход сопровождаемся освобождением потенциальной (свободной) энергии, запасенном и более упорядоченной структуре - спирали.
Эта энергия частично расходуется на тянущее усилие- перемещение нити актина на один шаг, а частично деградирует в тепло. Изменение конформации мостика одновременно вызывает сближение АТФ с АТФ-азным участком миозина, что вызывает гидролиз АТФ.
Часть освободившейся энергии рассеивается в виде тепла, а часть ее идет на восстановление спиральной конфигурации мостика, который выпрямляется по мере ресинтеза АТФ или поступления новых молекул АТФ извне. Актомиозиновый комплекс распадается и цикл может повториться, если в системе присутствуют ионы кальция.
При отсутствии в системе молекул АТФ она будет находиться в состоянии окоченения - молекулы актина будут оставаться присоединенными к связывающим центрам миозина.
При очень сильных мышечных сокращениях отмечается не только продвижение актиновых нитей, но и укорочение саркомеров в целом.
РАБОТА И СИЛА МЫШЦ
Величина сокращения (степень укорочения) мышцы при данной силе раздражения зависит как от ее морфологических свойств, так и от физиологического состояния. Длинные мышцы сокращаются на большую величину, чем короткие. Умеренное растяжение мышцы увеличивает ее сократительный эффект, при сильном же растяжении сокращение мышцы ослабляется. Если в результате длительной работы развивается утомление мышцы, то величина ее сокращения падает.
Для измерения силы мышцы определяют тот максимальный груз, который она в состоянии поднять. Эта сила может быть очень велика. Так, установлено, что собака мышцами челюсти может поднять груз, превышающий вес ее тела в 8,3 раза. О силе икроножных мышц человека судят по величине груза, положенного ему на плечи, с которым он в состоянии приподняться на носки.
Сила мышцы при прочих равных условиях зависит не от ее длины, а от поперечного сечения: чем больше физиологическое поперечное сечение мышцы, т. е. сумма поперечных сечений всех ее волокон, тем больше тот груз, который она в состоянии поднять. Физиологическое поперечное сечение совпадает с геометрическим только в мышцах с продольно расположенными волокнами; у мышц с косыми волокнами сумма поперечных сечений волокон может значительно превышать геометрическое поперечное сечение самой мышцы. По этой причине сила мышцы с косыми волокнами значительно больше, чем сила мышцы той же толщины, но с продольными волокнами. Чтобы иметь возможность сравнивать силу разных мышц, максимальный груз, который мышца в состоянии поднять, делят на число квадратных сантиметров ее физиологического поперечного сечения. Таким образом вычисляют абсолютную мышечную силу.
Большинство мышц млекопитающих и человека имеет перистое строение. Перистая мышца имеет большое физиологическое сечение, а поэтому обладает большой силой.
Работа мышцы измеряется произведением поднятого груза на величину укорочения мышцы, т. е. выражается в килограммометрах или граммсантиметрах.
Между грузом, который поднимает мышца, и выполняемой ею работой существует следующая зависимость. Внешняя работа мышцы равна нулю, если мышца сокращается без нагрузки. По мере увеличения груза работа сначала увеличивается, а затем постепенно падает. При очень большом грузе, который мышца не способна поднять, работа становится равной нулю.
Мощность мышцы, измеряемая величиной работы в единицу времени, также достигает максимальной величины при средних нагрузках. Поэтому зависимость работы и мощности от нагрузки получила название правила средних нагрузок.
УТОМЛЕНИЕ МЫШЦЫ
Утомлением называется временное понижение работоспособности клетки, органа или целого организма, наступающее в результате работы и исчезающее после отдыха.
Если длительно раздражать ритмическими электрическими стимулами изолированную мышцу, в которой подвешен небольшой груз, то амплитуда ее сокращений постепенно убывает, пока не дойдет до нуля. Полученная таким образом кривая называется кривой утомления. Измерив и суммировав высоту всех сокращений, можно узнать общую высоту подъема груза, а умножив груз на эту величину, определить количество работы, выполненной мышцей до наступления полного утомления.
Наряду с изменением амплитуды сокращений при утомлении нарастает латентный период сокращения и увеличиваются пороги раздражения и хронаксия, т. е. понижается возбудимость. Следует, однако, подчеркнуть, что все эти изменения возникают не тотчас же после начала работы мышцы - существует некоторый период, в течение которого наблюдаются увеличение амплитуд сокращений и небольшое повышение возбудимости мышцы. При этом мышца становится легко растяжимой. В таких случаях говорят, что мышца «врабатывается», т. е. приспособляется к работе при заданном ритме и силе раздражения. При дальнейшем, длительном раздражении наступает утомление мышечных волокон.
Эргография
Для изучения мышечного утомления у человека в лабораторных условиях пользуются эргографами - приборами для записи амплитуды движения, ритмически выполняемого группой мышц.
ЗАКЛЮЧЕНИЕ
Таким образом, можно сделать вывод, что мышца обладает способностью трансформировать энергию в механическую, производя при этом работу. Эта работа затрачивается на выполнение произвольных движений, а также на моторику внутренних органов. По своим свойствам мышцы отличаются от обычных твердых тел и относятся к эластомерам - материала типа каучука.
Сократительная система мышц состоит из сократительных и эластических элементов.
Химическая энергия мышцы превращается в механическую энергию сокращения без промежуточного превращения в тепло. Во время сокращения энергия затрачивается не только на работу, совершаемую мышцей, но и на выделения тепла. При работе теплопродукция мышц значительно увеличивается и находится в прямой зависимости от скорости сокращения мышц - при медленном сокращении в единицу времени тепла выделяется меньше, чем при быстром. Работа, производимая мышцей за единицу времени, т.е. мощность будет равна произведению напряжения на скорость сокращения.
Мощность мышцы зависит от нагрузки и скорости сокращения мышцы.
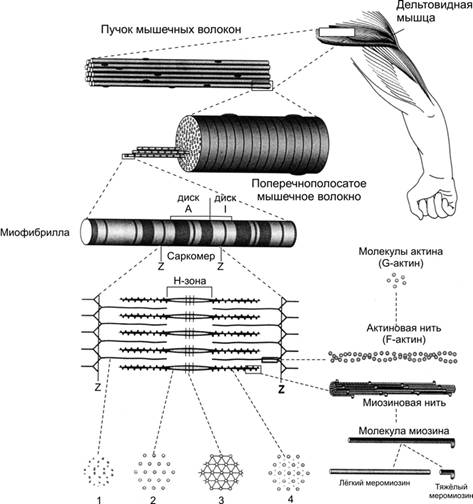
Каждое мышечное волокно представляет собой симпатическую многоядерную структуру. Мышечное волокно содержит миофибриллы, которые состоят из протофибрил. Одни нити образованы молекулами белка миозина, а другие молекулами белка актина.
В мышце миозин и актин способны образовывать комплексное соединение - актомиозин.
АТФ, образующаяся в процессах окисления и фосфорирования, является источником мышечного сокращения. В мышечном сокращении АТФ играет двойную роль: способствует диссоциации актомиозина на актин и миозин и одновременно под влиянием аденозитрифосфатазных свойств миозина сама расщепляется, освобождая энергию. Мышечное сокращение возникает в результате возбуждающего действия нервного импульса, проходящего в нервные окончания мионевральных синапсов.
В настоящее время более широкое распространение получила теория «скользящих нитей». Эта теория, разработанная Л.Хаксли, Дж.Хансон и М.Хакси, она заключается в том, что при мышечном сокращении тонкие актиновые нити продвигаются и скользят между толстыми миозиновыми нитями к центру саркомера.
Таким образом, сокращение мышц является процессом, иллюстрирующим сопряжения функции (энергетических процессов) и структуру (механизмов, участвующих в сокращении) живой клетки.
СПИСОК ЛИТЕРАТУРЫ
. Биофизика: Учеб. для вузов. / Под ред. П.К.Костюка - Киев: Выщэ. школэ 2008. - 504 с.
. Пирузян Л.А., Ландау М.А. Вопросы медицинской биофизики. 2009.
. Ремизов А.Н. Медицинская и биологическая физика: Учеб. для мед. вузов. 1987 - 638 с.
. Биофизика: Учеб. для студ. мед.ин-ов / Ю.А.Владимиров и др. - М.: Медицина, 2008. - 272 с.
. Волькенштейн М.В. Физика и биология. - М.: Наука, 2005. - 152 с.
. Губанов Н.И., Утепбергеров А.А. Мед. физика: Учеб. для мед.ин-ов. - М.: Медицина, 1978. - 335 с.
. Беликова З.П. и Павлова Р.С. Учебное пособие по мед. биофизике (Избр. главы). М.: Медицина, 1969.
. Биофизические методы исследования: Пер. с англ. (Под ред. Ф.Юбера - М.:1956 - 404 с.
. Ефимов В.В. Биофизика для врачей. М., МедГиз, 1952 - 342 с.
. Е.Б.Бабский, А.А.Зубков., Г.И.Косицкий, Б.И.Ходоров. «Физиология человека». 2008.
. Физиология человека. Под ред. Г.И.Косицкого. М., 2007.
. Основы физиологии человека (в 2-х томах). Под ред. Б.И.Ткаченко. СПб., 2004.
Date: 2016-05-23; view: 1368; Нарушение авторских прав