Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Экологический словарь - минимум 5 page
|
|
Факторы, влияющие на смертность. В основном смертность и возрастная структура видов зависят от того, какие шансы для выжива-ния имеют особи в различных возрастных группах. Наглядную информацию дает кривая выживаемости отдельных видов, полученная путем подсчета процента живых особей в отдельной популяции. Для большинства видов такая кривая обычно может быть одним из трех наиболее типичных вариантов (рис. 13).
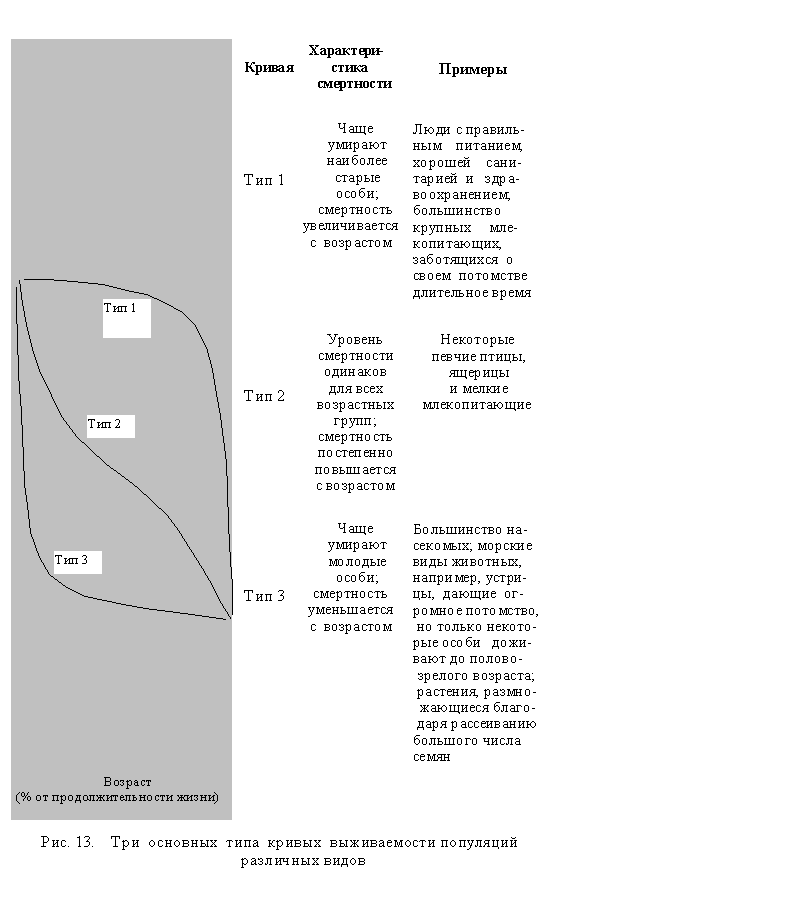
Смертность зависит и от других факторов. Один из них - межвидовая конкуренция: конкуренция между особями двух или более различных видов за питание и другие ресурсы. Другой фактор - это внутривидовая конкуренция: конкуренция между особями одного вида за скудные ресурсы. Смертность может также повыситься из-за нали-чия хищников, болезней и паразитов, стресса от перенаселенности, потери или ухудшения естественной среды обитания в результате человеческой деятельности и, кроме того, из-за природных катаклизмов, таких, как засухи, землетрясения, ураганы, пожары и наводнения.
Экспоненциальный рост продолжается вплоть до внезапного паде-ния плотности популяции в результате исчерпания ресурсов среды. Такой рост не зависит от плотности, так как его регуляция не связана с плотностью популяции до самого момента катастрофы. Едва ли не ка-ждый организм способен увеличить свою численность до заселения всей Земли при условии достатка пищи, воды, пространства и защиты от врагов. При неограниченных ресурсах такие быстроразмножающиеся виды как бактерии, насекомые, мыши и некоторые рыбы, могут сделать это за короткий отрезок времени. Например, при отсутствии ограничений один вид бактерий полностью заселил бы нашу планету за 30 дней. Почему же этого не происходит? Потому что природные условия не столь идеальны, а ресурсы ограниченны. Такие факторы, как наличие хищников, внутри- и межвидовая конкуренции, недостаток пищи, болезни, неблагоприятные климатические условия, отсутствие подходящих местообитаний, как правило, останавливают рост популяции ниже уровня максимальной рождаемости. Максимальный размер популя-ции одного вида, который природная экосистема способна поддерживать в определенных экологических условиях неопределенно долго, называется поддерживающей емкостью экосистемы для данного вида или просто емкостью экосистемы.
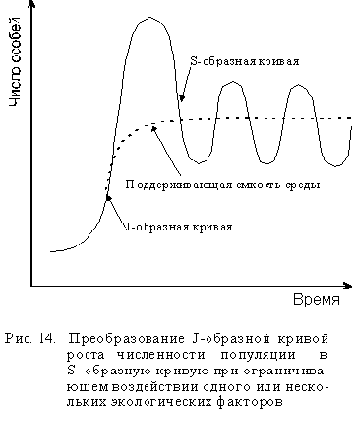
При ограниченных ресурсах размеры популяции того или иного вида также ограниченны и смертность начинает расти, когда числен-ность популяции достигает или временно превышает емкость экосистемы. Когда это случается, J-образная кривая роста популяции начинает плавно изгибаться и принимает вид S-образной кривой (см. рис. 14). Такой тип роста называют зависимым от плотности, так как скорость роста зависит от плотности популяции, которая влияет на истощение пищевых ресурсов и накопление токсичных продуктов, а потому и на рост. С увеличением плотности скорость роста популяции постепенно снижается до нуля, а кривая выходит на плато. При нулевом росте популяция стабильна, то есть размеры ее не меня-ются, при этом отдельные организмы растут и размножаются. Нулевая скорость роста популяции означает лишь то, что скорость размножения уравновешена смертностью. После этого размеры популяции обычно колеблются в пределах емкости среды.
Переход от J-образной к S-образной кривой может быть достаточно плавным или же может наблюдаться резкое падение численности популяции, называемое крахом или коллапсом популяции. Кол-лапс популяции возникает, когда численность размножающейся популяции превысит емкость экосистемы или когда изменения условий окружающей среды внезапно понижают емкость экосистемы. Тогда ог-ромное число особей, неспособных мигрировать в другие места, погибает.
Рост, соответствующий J- и S-образным кривым, можно описать алгебраически с помощью обыкновенных дифференциальных уравнений (оба уравнения относятся к популяциям, в которых поколения полностью перекрываются, так что популяция изменяется непрерывно; это и позволяет использовать дифференциальные уравнения).
Если обозначить N - число особей в популяции и t- время, то скорость изменения численности во времени dN/dt пропорциональна N, то есть

где r - константа, врожденная скорость роста численности популяции, связанная с максимальной скоростью размножения особей данного вида; чем выше скорость размножения, тем больше значение r. Уравнение (1) описывает J-образную кривую роста популяции, где рост не зависит от плотности. Если r положительно, численность популяции увеличивается экспоненциально; если r отрицательно, численность популяции уменьшается экспоненциально. Это отражает быстрые увеличения и уменьшения численности популяции. Размеры популяции при этом не стабилизируются.
Введем в уравнение (1) поддерживающую емкость среды К. Она отражает влияние среды на снижение роста численности до определенного стационарного уровня, то есть описывает S-образную кривую роста или роста, зависящего от плотности:

Если N > K, скорость роста отрицательна, если же K > N, ско-рость роста положительна и численность популяции стремится к K=N, то есть приводится в соответствие с поддерживающей емкостью среды. При K=N скорость роста популяции равна нулю и размеры попу-ляции остаются постоянными.
J- и S-образные кривые - это идеальные модели роста популяции. При их рассмотрении и описании предполагается, что все организмы сходны между собой, имеют равную способность к размножению и равную вероятность погибнуть, так что скорость роста в экспоненциальной фазе зависит только от ее численности и не ограничена условиями среды, которые остаются постоянными. Однако в отноше-нии природных популяций эти предположения часто неверны.
Модели динамики популяций в природе. Математическую модель динамики популяции, изображенную на рис. 14, когда J-образная кривая преобразуется в S-образную кривую можно наблюдать лишь в лабораторных условиях. В природе, однако, мы встречаемся с тремя основными типами кривых роста численности популяций: относительно стабильного, скачкообразного и циклического (рис. 15).
Виды, чья численность находится из года в год на уровне емкости окружающей среды, имеют относительно стабильные популяции. Подобное постоянство в размерах популяции характерно для многих видов дикой природы и встречается, например, в нетронутых тропических дождевых лесах, где среднегодовая температура и количество осадков крайне мало изменяются день ото дня и год от года.
Некоторые виды, такие, как енот, обычно имеют довольно ста-бильную численность популяции, но иногда их число резко возраста-ет, или "подскакивает" до наивысшей отметки, а затем стремительно падает до какогото относительно стабильного низкого уровня. Такие виды относятся к популяциям со скачкообразным ростом численно-сти (см. рис. 15).
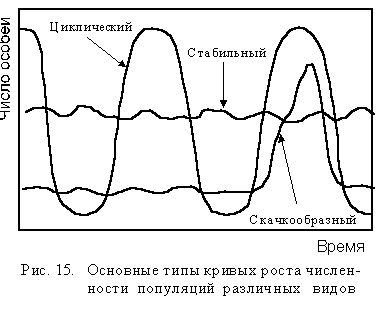
Внезапные увеличения численности происходят тогда, когда временно повышается емкость среды для данной популяции, что может быть связано с улучшением погодных условий и питания или с резким уменьшением численности хищников, врагов и т.п. После того, как численность популяции превысит эту новую, более высокую емкость системы, смертность в популяции резко увеличивается и размеры популяции значительно сокращаются.
Некоторые виды каждые три-четыре года или каждые десять лет, или в иные постоянные временные интервалы резко увеличивают число особей, что затем сопровождается таким же резким падением численности популяции. Данные виды относятся к популяциям с циклическим ростом их численности (см. рис. 15). Фактор или группа факторов, влияющих на подобную динамику численности, пока еще недостаточно изучены.
5.3. Принцип стабильности экосистем
Все рассмотренные в предыдущем разделе закономерности ка-саются одной отдельно взятой популяции. В природных экосистемах популяции всех видов, населяющие данный биотоп, находятся в непрерывном взаимодействии между собой, образуя биотическое сообщество.
Когда экосистема включает небольшое число видов, пищевые связи в ней достаточно просты и очевидны: например, хищник - жертва, паразит - хозяин, фитофаг - растение и т.п. Для такого типа отно-шений в изменении численности взаимодействующих видов характерен сдвиг по фазе j (рис 16):
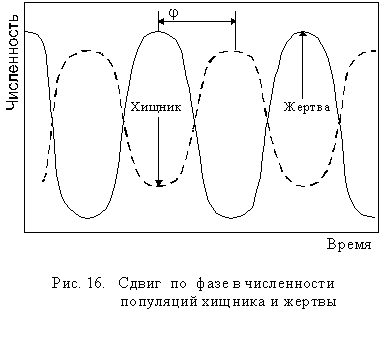
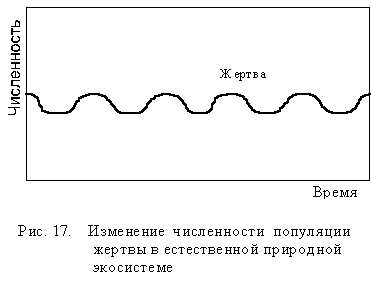
Однако в экосистеме каждый организм зависит одновременно от нескольких хищников, нескольких паразитов, которых всех вместе называют естественными врагами. Поэтому в естественных экосистемах речь идет о равновесии между организмами данного вида и его естественными врагами. В единой пищевой сети равновесие значительно устойчивее и менее подвержено резким колебаниям, так как разные враги начинают снижать численность вида при разной плотности его популяции. Это значительно ослабляет изменение популяции жертвы (рис 17).
Динамическое равновесие популяций в экосистеме не возникает автоматически, а устанавливается на протяжении многих тысяч и даже миллионов лет. За это время виды адаптируются друг к другу и к среде своего обитания, так что естественные враги поэтому сами останутся без источника существования. Они лишь ограничивают рост популяции при повышении ее плотности. Аналогичным образом у видов есть определенная устойчивость к естественным паразитам и болезнетворным агентам, которые не приводят к полному вымиранию популяции, но также регулируют ее плотность.
Такое взаимоприспособление крайне важно для равновесия популяций, а следовательно, и экосистем в целом. Популяции, разви-вающиеся в изоляции друг от друга, как правило, не могут существо-вать в равновесии. Например, интродуцированный вид не всегда сталкивается с естественными врагами, в результате чего происходит популяционный взрыв его численности или, наоборот, он оказывается слишком "эффективным" естественным врагом, уничтожающим другие виды.
5.4. Реакция популяций на постепенное изменение условий окружающей среды
Итак, видовое разнообразие сообщества обеспечивает стабиль-ность экосистем. Равновесие экосистемы определяется равновесием составляющих ее популяций. Это динамическое равновесие складыва-ется в процессе длительного взаимодействия популяций и является результатом адаптации популяций друг к другу.
Рассмотрим, как реагирует та или иная популяция на изменение условий окружающей среды.
Существуют два принципиально разных типа изменений условий окружающей среды: медленное, постепенное, не приводящее к нарушению равновесия в экосистеме, и стрессовое, вызывающее нарушение равновесия вплоть до разрушения и уничтожения экосистемы.
Рассмотрим сначала реакцию популяций на изменения первого типа. Она может быть двоякой: либо происходит биологическая эволюция данного вида, либо из данного вида образуются два или более новых. В основе того и другого процессов эволюции и видообразования лежит естественный отбор.
Естественный отбор и эволюция. Популяции многих видов приспосабливаются к изменению условий окружающей среды путем изменения своего генетического состава. У разных особей одного вида гены неидентичны. Эти генетические разнообразия помогают приспособиться и сохраниться многим видам.
Процесс, посредством которого определенные гены и комбинации генов в популяциях воспроизводятся чаще других, называется естественным отбором.. Чарльз Дарвин, впервые выдвинувший эту идею в 1858 году, определил естественный отбор как выживание наиболее приспособленных. Это означает, что выживают особи с наиболее приспособленными к существующим условиям окружаю-щей среды генетическими свойствами, в среднем дающие большее потомство в следующем поколении.
Виды, которые часто дают многочисленное мелкое потомство с короткой продолжительностью жизни, могут приспосо-биться к изменениям условий окружающей среды путем естественного отбора за очень короткий срок, как это делают сорные травы, насекомые, грызуны, бактерии.
Другие виды, такие, как слоны, тигры, акулы, человек, имеют длительный период воспроизводства и малочисленное потомство. У таких видов адаптация к изменениям окружающей среды занимает обычно тысячи и миллионы лет.
Окончательные изменения в генетической структуре попу-ляции, вызванные переменой условий окружающей среды и происшедшие в силу разной способности к размножению и естественного отбора, называются биологической эволюцией или просто эволюцией. Рассмотрим механизм эволюции.
До сих пор строгого, точного и однозначного ответа на вопрос: "что такое жизнь?" никто дать не может. Однако всякое живое вещество всегда и везде обладает по крайней мере тремя свойствами: 1) воспроизводимостью, то есть способностью порождать себе подобное; 2) н аследственностью - способностью передавать родительские черты детям; 3) изменчивостью, то есть способностью к изменению - мутации.
Роль этих факторов в живой природе неоценима. Без воспроизведения жизнь перестала бы существовать. Без наследственности не было бы преемственности между поколениями и, значит, видовые особенности родителей не передавались бы детям. Наконец, без мутации не было бы изменчивости и развитие жизни не пошло бы дальше ее первоначальных форм.
Напомним кратко механизм образования мутаций, так как они лежат в основе более общего механизма эволюции живого вещества.
Ткани любого живого организма состоят из клеток. В ядре каждой клетки находятся нитевидные хромосомы, представляющие собой гигантские молекулы ДНК (их общая протяженность в организме человека достигает до 1011 км) в соединении с молекулами белков. Отдельные участки хромосомы, ответственные за те или иные наследственные изменения, называются генами. Различный порядок сле-дования генов в молекулах ДНК кодирует различные свойства будущего организма - его генотип. Генотип каждого индивидуума уникален, что и определяет его непо-хожесть на других (идентичный генотип в природе имеют только однояйцевые близнецы).
При делении клетки сначала делятся хромосомы: каждая из них удваивается и образует совершенно одинаковые хромосомы, которые тут же расходятся. После деления хромосом делится ядро, а затем остальное вещество клетки и обе половины оформляются в самостоятельные клетки. Из одной клетки возникают сначала 2 со-вершенно одинаковых, затем 4, 8, 16 и т.д.
Процесс удвоения хромосом чрезвычайно строг, это исключительно точный и безошибочный механизм: при зарождении организма из одной клетки развива-ются миллионы с совершенно одинаковыми наборами хромосом. Однако в мире нет ничего абсолютного, имеется предел точности и у такого процесса. Иногда в этом процессе что-то нарушается - происходит случайный сбой, "опечатка" при копировании хромосомы и наследственная информация, которую несет хромосома, случайно становится "чуть-чуть" иной. Процесс случайного изменения хромосомы получил название мутации.
Причины природной мутации могут быть самые разные: тепловые колебания молекул, радиация, температура, химические воздейст-вия и просто случайные помехи в процессе удвоения хромосом. Когда "мутированная" хромосома удваивается, она повторяет структуру, полученную в результате мутации. Следовательно, наследство мутированной хромосомы также мутировано. А далее действует принцип усиления. По мере развития организма происходят многократные де-ления клеток и в итоге генмутант оказывается продублированным примерно 1015 раз. Таким образом, в процессе становления живого организма случайные изменения генетической наследственной програм-мы оказываются многократно усиленными. Тем самым случайные из-менения на атомномолекулярном уровне переходят на макроуровень - фенотип. Фенотип - это совокупность внутренних и внешних признаков организма: анатомических, физиологических, психологических, поведенческих и т.п. На этом уровне вступает в действие механизм естественного отбора живых организмов. Закон естественного отбора действует в природе очень жестко: уничтожает те организмы, которые в результате случайной мутации оказались неприспособленными, а из тех, которые оказались в той или иной мере приспособленными к ус-ловиям обитания, отдает предпочтение более приспособленным, а менее приспособленных, как правило, также уничтожает. Природа ни о ком и ни о чем не заботится. Все дело в жестком принципе отбора наиболее приспособленных; иной мудрости, иной стратегии у природы нет.
Таким образом, если мутации обеспечивают случайное отклонение организма от исходного состояния, то естественный отбор как бы оценивает результат этого отклонения. Случайные мутации в соче-тании с естественным отбором представляют собой движущую силу эволюции.
С точки зрения эволюции мутации, безусловно, полезны. Более того, они необходимы. Огромное разнообразие генов у каждого вида, а также многообразие существующих на Земле видов - все это следствие многочисленных мутаций, которые происходили на протяжении многих миллионов лет и происходят поныне.
С точки зрения отдельных организмов мутации, как правило, вредны, в отдельных случаях даже смертельны. Причина этого заключается в том, что каждый организм есть результат длительной эволю-ции, он чрезвычайно тонко приспособлен к окружающей среде и далеко не каждое изменение его структуры, вызванное мутацией, идет ему впрок. Скорее наоборот. Для улучшения высокоорганизованного организма нужны мутации специального характера, которые встречаются крайне редко, и может пройти много времени, прежде чем нужная с точки зрения приспосабливаемости новая мутация будет найдена. Может случиться так, что за время, пока она произойдет, организмы этого вида вымрут, причем не столько из-за отсутствия нужной мутации, сколько из-за избытка ненужных и вредных, воздействующих на жизненно важные органы и функции организма и снижающих или даже разрушающих его приспособленность, имеющуюся ранее.
Таким образом, случайные мутации столь же вредны, как и необходимы. Вид, у которого мутации возникают слишком часто, на-пример в результате радиоактивного заражения или химического за-грязнения среды обитания, может исчезнуть потому, что многие его представителимутанты станут неприспособленными к окружающей среде. Если, напротив, у данного вида мутации происходят слишком редко, то при какомнибудь значительном изменении внешних условий у вида не окажется необходимого запаса изменчивости, он не сможет быстро приспособиться и также погибнет. По одной из гипо-тез так, по-видимому, относительно недавно вымерли мамонты, не сумевшие приспособиться к быстрым изменениям климата на Земле во время ледникового периода.
Видообразование. Сейчас на Земле существуют около 5 млн ви-дов живых организмов, около 1,5 млн из них известны ученым и опи-саны. Виды возникли в результате взаимодействия на протяжении миллиардов лет двух процессов. Один из них - образование видов. При изменении условий окружающей среды в результате внутреннего отбора возникают новые виды, хорошо приспособленные к этим условиям. Другой процесс - вымирание. В изменившихся условиях при-родной среды многие виды, не сумевшие генетически к ним адаптиро-ваться и нормально размножаться, перестают существовать.
Биологи считают, что примерно 94 - 99 % когда-либо существовавших на Земле видов вымерли. Однако тот факт, что сейчас на Земле насчитывается около 5 млн видов (по другим оцен-кам около 30 млн) расценивается как преобладание процесса образования видов над вымиранием.
Видообразование может происходить несколькими путями:
1) в результате изменения в одной генетической линии;
2) путем постепенного расхождения линий потомков на два и более видов в ответ на изменение условий среды. Такой путь видообразования характерен для тех случаев, когда популяции одних и тех же видов распространились по ареалам с различным климатом, источниками питания, почвами и т.п. и остались там на длительное время, обычно время жизни от 1000 до 100000 поколений, например, в результате геологического раскола одного участка суши на два раздельных острова, изменения русла реки, поднятия горного массива;
3) в результате миграции части группы, например, в поисках пищи.
Со временем изолированные группы популяций, находясь в различных условиях местообитания, начинают вследствие естественного отбора генетически сильно отличаться. При достаточно длительной изоляции в совершенно разных условиях особи двух разных популя-ций, даже если займут один и тот же ареал, уже не будут способны к скрещиванию. Таким образом, из одного вида образовались два раз-ных вида. У многих быстро размножающихся организмов видообразование может занять тысячи или даже сотни лет, однако в большинстве случаев на это требуется от десятков тысяч до миллионов лет.
Биологическое разнообразие. В результате естественного отбора за миллиарды лет появился самый ценный "ресурс" планеты - биологическое разнообразие (биоразнообразие). Оно включает в себя два взаимосвязанных понятия: генетическое разнообразие и видовое разнообразие. Генетическое разнообразие - это многообразие генетических свойств у особей одного вида. Видовое разнообразие - это число различных видов внутри какоголибо сообщества организмов.
Невероятное генетическое разнообразие на нашей планете позволяет видам непрерывно изменяться. Каждый ныне существующий вид представляет собой накопленную генетическую информацию, которая позволяет ему приспособиться к определенным изменениям окружающей среды. Биоразнообразие - это "страховая политика" природы против катастроф. Сохранение генофонда планеты, а значит и биоразнообразия, одна из важнейших задач человечества.
5.5. Изменение экосистем при стрессовых воздействиях. Экологические сукцессии
Крупномасштабные стрессовые воздействия выводят экосистему из состояния равновесия, что часто приводит к ее разрушению. В ре-зультате естественных катаклизмов (пожаров, наводнений, изверже-ний вулканов, землетрясений, отступания ледников) или антропогенных изменений окружающей среды (пожаров, открытой добычи полезных ископаемых, загрязнения, затопления участков суши под водохранилища, расчистки территорий под пашни) существовавшая ранее на данной территории экосистема разрушается.
После такого крупномасштабного нарушения " пострадавший" участок начинает возрождаться в несколько этапов.
Сначала несколько неприхотливых пионерных видов заселяют территорию и начинают формировать почвы, а в водных экосистемах - донные отложения. В конце концов эти пионерные виды изменяют почву (или отложения) и прочие условия настолько, что участок ста-новится менее пригодным для них самих и более пригодным для но-вой группы растений и животных с другими экологическими требова-ниями. Такой процесс, при котором сообщества видов растений и животных замещаются с течением времени другими, обычно более сложными сообществами, называется экологической сукцессией или просто сукцессией. Экологическая сукцессия обычно про-должается до тех пор, пока сообщество не станет стабильным и само-обеспечивающимся. Конечное сообщество, занявшее нарушенный участок, называется зрелым сообществом, а экосистема в целом - климаксовой. В разных абиотических условиях формируются неоди-наковые конечные климаксовые экосистемы. В сухом и жарком климате это будет пустыня; в жарком, но влажном - тропический лес и т.п. На планете выделяют несколько крупных наземных климаксовых экосистем, которые называют биомами: тундра, хвойные леса (тайга), леса умеренного пояса, степи, пустыни, саванны, дождевые тропиче-ские леса. Таким образом, основные б иомы Земли - это климаксовые экосистемы соответствующих географических зон.
Экологи выделяют два вида экологических сукцессий: первичные и вторичные. Какой из типов наблюдается - зависит от условий участка в начале процесса.
Первичная сукцессия - это последовательное развитие сооб-ществ на участках, лишенных почв. Такими участками могут быть скалы, глины после отступания ледника или прохождения селя, остывшая лава, участки открытой добычи полезных ископаемых, с которых снят верхний слой почвы и т.п. На таких бесплодных участках первичная сукцессия от голой скальной породы до зрелого леса может занять от сотен до тысяч лет.
Более распространенным типом сукцессий является вторичная сукцессия, то есть последовательное развитие сообществ в ареале, в котором естественная растительность была устранена или сильно нарушена, но почва не была уничтожена. Участками, на которых может наблюдаться вторичная сукцессия, могут быть заброшенные сельхозугодья, сгоревшие или вырубленные лесные участки, земли, где растительность уничтожена в результате ведения горностроительных работ, сильно загрязненные водные источники, затопленные ранее территории и т.п. Поскольку имеется в наличии какой-то слой почвы, новая растительность может появиться в течение всего нескольких недель.
Вновь созданные озера, водохранилища и пруды также пережи-вают вторичную сукцессию. По мере их постепенного заполнения донными отложениями они в конечном итоге превращаются в назем-ные экосистемы, которые затем проходят вторичную наземную эколо-гическую сукцессию.
В уравнениях (1) и (2), описывающих рост популяции, были ис-пользованы параметры r и К. Виды, которые быстро размножаются, имеют высокое значение r и называются r-видами. Это обычно "оппортунистические" виды - типичные пионерные виды нарушен-ных местообитаний. Такие местообитания называют r-отбирающими, так как они благоприятствуют росту численности r-видов.
Виды с относительно низким значением r называют К-ви-дами. Скорость их размножения чувствительна к плотности популяции и остается близкой к уровню равновесия, определяемому величи-ной К. Они более характерны для поздних стадий сукцессии. Об этих двух типах видов говорят, что они используют соответственно r-стратегию и К-стратегию. Хотя существует и целый спектр промежуточных стратегий, эти две стратегии имеют важное значение в попу-ляционной экологии и в экологии сообществ. Некоторые характерные особенности r- и К-стратегий приведены в табл. 4.

По существу эти две стратегии представляют два различных ре-шения одной задачи - задачи длительного выживания вида. Виды с r-стратегией быстрее заселяют нарушенные местообитания, характер-ные для ранних стадий сукцессий, такие, как обнаженная горная по-рода, лесные вырубки, выгоревшие участки, чем виды с К-стратегией, так как они легче распространяются и быстрее размножаются. Виды с К-стратегией более конкурентоспособны, и в конце концов они вытесняют r-виды, которые тем временем перемещаются в другие нарушенные местообитания. Высокий репродуктивный потенциал r-видов оз-начает, что, оставшись в каком-либо местообитании, они быстро использовали бы доступные ресурсы и превысили поддерживающую емкость среды, а затем популяция погибла бы. Другими словами, для них характерна J-образная кривая роста с быстрым падением численности популяции в конце. Виды с r-стратегией занимают данное местообитание лишь в течение жизни одного или самое большее нескольких поколений. Затем они переселяются на новое место. Отдельные попу-ляции могут регулярно вымирать, но вид при этом перемещается и выживает. В целом эту стратегию можно охарактеризовать как страте-гию "борьбы или бегства". Хотя для самых мелких организмов, таких, как бактерии и насекомые, характерна r-стра-тегия, а для самых крупных, таких, как позвоночные и деревья, - К-стратегия, большинству организмов свойственны стратегии промежуточных типов.
Сукцессия любого масштаба заканчивается формированием зрелого сообщества, и в экосистеме все популяции приходят в состояние динамического равновесия. Далее эти экосистемы существуют в соответствии с рассмотренными ранее законами и принципами функционирования экосистем.
Сукцессии любого масштаба характеризуются рядом общих закономерностей, многие из которых важны для практической деятельности человека.
В ходе сукцессии постепенно нарастает видовое разнообразие. Это ведет к усложнению связей внутри сообщества, разветвлению цепей питания и усложнению трофической сети, усилению регуляторных возможностей внутри системы, ее стабильности.
Серьезные преобразования происходят в энергетическом балансе системы (рис. 18).
С энергетических позиций сукцессия - это такое неустойчивое состояние сообщества, которое характеризуется несоответствием двух показателей: валовой продуктивности и энергетических затрат всей системы на поддержание жизнедеятельности - дыхания.
В ходе сукцессии общая биомасса сообщества Б сначала возрастает, но затем темпы этого роста снижаются и на стадии климакса биомасса системы стабилизируется.
На первых этапах сукцессии чистая продуктивность сообщества Пч относительно высока. В зрелых, устойчивых экосистемах практически весь прирост растительности поступает и расходуется в цепях питания гетеротрофами, поэтому чистая продуктивность сообщества приближается к нулю.
Date: 2016-06-08; view: 784; Нарушение авторских прав