Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Тема «Гаструляция у зародышей беззубки и амфибии
Содержание
1. Гаметогенез 4
2. Половые клетки. Оплодотворение 13
3. Дробление 20
4. Гаструляция у зародышей беззубки и амфибии. Нейруляция 26
5. Гаструляция у рыб и птиц 37
6. Внезародышевые (провизорные) органы 53
7. Развитие млекопитающих 67
Тема «Гаметогенез»
Вопросы для самоконтроля:
1. Сперматогенез. Особенности этапов сперматогенеза. Роль клеток Сертоли.
2. Оогенез. Особенности этапов оогенеза.
3. Характеристика малого и большого роста ооцитов.
4. Способы питания ооцитов.
5. Блок мейоза.
Сравнительные особенности спермато- и оогенеза.
Препарат №1. Семенник крысы (рис. 1)
Гистологический срез, окраска гематоксилином и эозином.
Данный препарат представляет собой срез семенника крысы.
Семенник - мужская половая железа, в которой развиваются мужские половые клетки.
Основной структурный элемент семенника - извитой каналец, который выстилается соматическими по происхождению клетками Сертоли (клетки-кормилки), включает в себя несколько клеточных слоев сперматогенного эпителия, представляющего собой дифференцирующиеся половые клетки, развитие которых происходит в направлении от базальной мембраны (базальная зона) к просвету канальца (адлюминальная зона). На срезе канальцы выглядят округлыми либо овальными в зависимости от плоскости сечения.
Основания клеток Сертоли находятся на базальной мембране канальца, а ветвящиеся отростки обращены в сторону его просвета и образуют специфические контакты с клетками сперматогенного эпителия. Рассмотреть границы клеток Сертоли невозможно, но хорошо различимы ядра. Обширная светло-розовая цитоплазма заполняет все промежутки между половыми клетками. Светлые ядра клеток Сертоли, обычно треугольной либо овальной формы, содержат одно ядрышко, располагаются в базальной части клетки. Функции их многообразны: трофическая, опорная, регуляторная, фагоцитоз. У взрослых особей эти клетки не делятся.
Сперматогенез начинается в периферических участках извитых 
Рис. 1. Поперечный срез извитого канальца семенника крысы.
канальцев. Между основаниями клеток Сертоли у базальной мембраны находятся клетки с относительно крупным ядром и небольшим количеством цитоплазмы — сперматогонии. Можно заметить различия в строении ядер сперматогоний разных генераций. Для сперматогониев типа А характерны два варианта ядер: с деконденсированным, равномерно распределенным хроматином (светлые ядра) и с конденсированным хроматином (темные ядра). Вблизи ядерной мембраны расположены ядрышки. В сперматогониях типа Б хроматин конденсирован и распределен у ядерной мембраны.
В базальной зоне расположены сперматогонии, находящиеся в состоянии митотического деления. Это зона размножения.
В адлюминальной зоне ярусами располагаются последовательно сперматоциты первого и второго порядков (зона роста и созревания) и ближе к просвету — сперматиды (зона формирования сперматозоидов).
Половые клетки, находящиеся на определенном этапе сперматогенеза, можно различить по характерным морфологическим особенностям. Сперматоциты первого порядка (сперматоциты 1) — это наиболее крупные по размерам клетки; их ядра почти в два раза больше ядер сперматогоний и имеют характерную сетчатую структуру ядер, которая особенно характерна для пахитены и диплотены профазы мейоза.
По завершении ядерных перестроек в периоде роста сперматоциты 1 приступают к делениям созревания. В результате первого (редукционного) деления образуются сперматоциты второго порядка (сперматоциты 2). Они меньше по объему и располагаются ближе к просвету канальца. Из-за краткости соответствующей интерфазы они крайне редко обнаруживаются на срезах, также имеют ядро с сетчатой структурой, но рисунок сетки менее «грубый», чем в ядрах сперматоцитов 1.
Самый верхний ярус стенки канальца занимают гаплоидные сперматиды, образующиеся после второго (эквационного) деления мейоза. Популяции сперматид, видимые на разных срезах семенных канальцев, существенно различаются, так как находятся на разных этапах трансформации сперматид в сперматозоиды. Как правило, на одном срезе стенки семенного канальца можно увидеть сперматиды сразу двух возрастов — ранние и поздние.
Ранние сперматиды — небольшие овальные клетки с хорошо заметными контурами светлых ядер, хроматин в которых дисперсно распределен и имеет тонкозернистую структуру. Очень часто на периферии ядра видна довольно крупная темная формирующаяся акросома сперматозоида.
В процессе спермиогенеза сперматида приобретает морфологические свойства, характерные для спермия: ядро уплотняется и образует основную часть головки, формируется акросома, из дистальной центриоли начинает расти жгутик, цитоплазма в основном редуцируется.
Поздние сперматиды имеют темную окраску ядер, их жгутики направлены в просвет канальца.
Картина на поперечных срезах в разных участках извитых семенных канальцев оказывается неодинаковой, что связано с этапом сперматогенеза в конкретном участке канальца. В некоторых случаях на срезе могут присутствовать сперматиды только одного возраста, полностью сформированные сперматозоиды в просвете канальцев также видны не на всех срезах. На завершающем этапе клетки обособляются от клеток Сертоли и сперматозоиды выходят в просвет канальца.
Между извитыми канальцами семенника располагаются соединительная ткань и клетки Лейдига - интерстициальные эндокриноциты, основная функция которых — синтез мужского полового гормона тестостерона, необходимого для осуществления сперматогенеза.
При работе с препаратом необходимо на малом увеличении найти поперечные срезы извитых канальцев семенника с разными этапами сперматогенеза. На большом увеличении следует рассмотреть различные стадии развития сперматозоидов (сперматогогии, стерматоциты 1 и 2, сперматиды, сперматозоиды) и клетки Сертоли.
Препарат №2. Яичник кошки (рис. 2)
Гистологический срез, окраска гематоксилином и эозином.
На препарате представлен поперечный срез яичника кошки – женской половой железы.
На поперечных срезах яичника различимы корковое вещество, в котором находятся фолликулы всех стадий развития, и мозговое вещество, в котором проходят крупные сосуды яичника. Снаружи железу покрывает однослойный кубический эпителий, который называют зачатковым, хотя у млекопитающих он не выполняет функцию формирования новых половых клеток. Под зачатковым эпителием находится тонкий слой белковой оболочки, далее – корковое вещество с многочисленными формирующимися фолликулами.
Развитие женских половых клеток млекопитающих осуществляется при активном участии фолликулярных клеток.
У млекопитающих первый этап оогенеза — размножение оогоний — происходит в эмбриональный период. При формировании яичника группа оогониев окружается соединительно-тканными клетками. Такая структура носит название
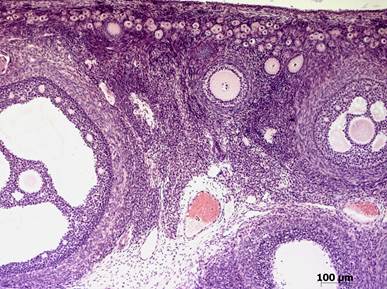
Рис. 2. Поперечный срез яичника кошки.
Пфлюгеровские шары. Позже, когда овогоний вступает в период малого роста (медленная стадия) происходит разделение структуры на отдельные фолликулы, представляющие собой совокупность одного овогония и окружающего его одного слоя фолликулярных клеток, преобразованных из соединительно-тканных. Возникает примордиальный фолликул. Располагаются примордиальные фолликулы, как правило, группами в самых поверхностных участках корковой области яичника, непосредственно под белковой оболочкой (генерация покоящихся фолликулов).
Примордиальные фолликулы порциями вступают в рост в течение всей жизни самки и оогонии превращаются в ооциты первого порядка, в ядре которых начинается профаза мейоза, но не завершается, а блокируется на стадии диплотены. Завершение профазы мейоза и дальнейшее развитие происходят лишь при половом созревании.
При инициации развития фолликулярные клетки начинают пролиферировать, из плоских превращаются в кубические, а затем в призматические и покрывают ооцит первого порядка сначала одним слоем (первичный фолликул), затем двумя, тремя и т.д. (вторичные фолликулы).
Первичный фолликул - ооцит первого порядка на стадии диплотены в окружении одного слоя призматических фолликулярных клеток. Между ними при совместном участии ооцита и окружающих его фолликулярных клеток начинает формироваться блестящая оболочка. Ее наружная часть, состоящая из кислых мукополисахаридов, секретируется фолликулярными клетками, а внутренняя, содержащая нейтральные мукополисахариды, образуется ооцитом первого порядка. Эту оболочку пронизывают микроворсинки ооцита и фолликулярных клеток.
Первичный фолликул увеличивается в объеме, как за счет увеличения размера самого ооцита, так и за счет увеличения числа клеток фолликулярного окружения, которые активно делятся в течение всего периода роста фолликула. При увеличении количества рядов фолликулярных клеток, окружающих овоцит первого порядка, более, чем на один, фолликул получает название вторичного фолликула.
Фолликулярная оболочка многослойного фолликула называется зернистой, прилежащие к ооциту фолликулярные клетки расположены радиально иобразуют вокруг ооцита лучистый венец, снаружи от зернистой оболочки начинает формироваться фиброзная оболочка.
Дальнейшее развитие фолликула заключается в появлении между фолликулярными клетками полостей, заполненных фолликулярной жидкостью и имеющих тенденцию к слиянию. Ооцит, находящийся в нем, все еще остается на стадии диплотены профазы мейоза. До тех пор пока продолжается рост самого ооцита первого порядка, фолликул остается компактным. Затем рост ооцита прекращается, а фолликул продолжает активно увеличиваться за счет пролиферации клеток зернистой оболочки и образования полостей. При слиянии полостей в общую центральную полость образуется третичный фолликул, или граафов пузырек. Это завершающий этап фолликулогенеза.
Зрелый фолликул состоит из нескольких клеточных слоев. Ооцит расположен среди поддерживающих его фолликулярных клеток, выдающихся в полости фолликула. Эта структура носит название яйценосного бугорка.
Ооцит окружен блестящей оболочкой и лучистым венцом. Полость фолликула ограничена зернистой оболочкой.Снаружи фолликул покрыт тонкой базальной мембраной и соединительно-тканной оболочкой (theca), состоящей из внутренней части (theca interna), содержащей сосуды, и наружной (theca externa), в состав которой входят стромальные и мышечные клетки.
Фолликулярные клетки играют роль не только опорных, ограничительных, трофических элементов, но вместе с текальными элементами являются железой внутренней секреции, вырабатывающей эстроген (женский половой гормон), заполняющий полость граафова пузырька. Кроме того, на них расположены рецепторы к гонадотропным гормонам гипофиза, под воздействием которых осуществляются развитие фолликулов и оогенез.
Если срез через графов пузырек прошел касательно, на препарате могут быть видны либо только стенки оболочки, либо оболочка и частично яйценосный бугорок.
Граафов пузырек на завершающих стадиях формирования быстро увеличивается в размерах, приближается к поверхности яичника и его верхушка (стигма) выдается над ней. Последующие события связаны с разрывом стенки фолликула в области стигмы, и выбросом фолликулярной жидкости вместе с ооцитом, окруженным блестящей оболочкой и фолликулярными клетками лучистого венца. Процесс разрыва фолликула и освобождения ооцита называется овуляцией. У большинства млекопитающих она происходит на стадии метафазы второго деления созревания ооцита. Завершение второго деления созревания, при котором образуются яйцеклетка и второе редукционное тельце, происходит лишь в процессе оплодотворения.
При работе с препаратом необходимо на малом увеличении найти примордиальные, первичные, вторичные и третичные (графов пузырек) фолликулы. При большом увеличении рассмотреть строение обозначенных структур.
Тема «Половые клетки. Оплодотворение»
Вопросы для самоконтроля:
1. Строение зрелой яйцеклетки.
2. Классификация яйцеклеток в зависимости от количества и распределения в них трофического материала.
3. Строение зрелого сперматозоида.
4. Биологическое значение оплодотворения.
5. Дистантные взаимодействия гамет при оплодотворении.
6. Контактные взаимодействия гамет при оплодотворении. Медленный и быстрый блоки полиспермии.
7. Ранние и поздние реакции яйцеклетки при оплодотворении.
8. Изменения, происходящие после проникновения сперматозоида в яйцеклетку.
9. Значение ооплазматической сегрегации.
10. Моно- и полиспермия.
Препарат № 1. «Изолецитальная яйцеклетка беззубки» (рис. 3).
Гистологический срез, окраска гематоксилином и эозином.
На срезе тела двустворчатого моллюска - беззубки - видны многочисленные срезы яичника и яйцеводов. Стенки яичника выстланы столбчатым или кубическим эпителием. Большинство срезов яичника имеют небольшой просвет и относительно толстую стенку, образованную желточными клетками цилиндрической формы с маленьким компактным ядром и цитоплазмой розового цвета. Среди этих клеток находятся базофильные клетки – яйцеклетки, на ранних этапах развития. В период роста яйцеклетки увеличиваются в размерах, приобретают оксифилию и перемещаются в просвет железы.

Рис. 3. Изолецитальная яйцеклетка беззубки. Поперечные срезы яйцеводов и яичников беззубки.
В начале малого роста яйцеклетки мелкие, содержат относительно крупное, бледно окрашенное ядро и 1-2 ядрышка. По мере роста в цитоплазме клеток накапливается РНК, увеличивается количество органелл, вследствие чего нарастает базофилия цитоплазмы и она приобретает красновато-фиолетовый цвет. В фазе большого роста яйцеклетка увеличивается в размерах, продвигается к просвету яичника. Вследствие интенсивного синтеза белков и желтка цитоплазма приобретает оксифильные свойства. Зрелые яйцеклетки теряют связь со стенкой яичника и оказываются в его просвете.
Срезы яйцеводов имеют больший просвет и относительно тонкие стенки, выстланные кубическими клетками с мелкозернистой цитоплазмой и мелкими компактными ядрами, среди них не встречаются созревающие яйцеклетки. В просвете яйцеводов свободно располагаются зрелые яйцеклетки.
Яйцеклетки беззубки олиголецитальные, изолецитальные, имеют округлую форму. Бледное ядро расположено в центре или несколько эксцентрично, содержит от одного до трех ядрышек.
На малом увеличении необходимо найти срезы яйцеводов и яичника, научиться их идентификации. На большом увеличении следует рассмотреть отличия в яйцеклетках разных возрастов.
Препарат № 2. «Умеренно телолецитальная яйцеклетка лягушки» (рис. 4).
Гистологический срез, окраска гематоксилином и эозином.
На препарате представлен срез кладки лягушки, в котором видны яйцеклетки разной степени зрелости, находящиеся на разных этапах стадии роста, имеющие поэтому разную величину и окраску.
В начале стадии роста яйцеклетки (оогонии) небольшого размера, их цитоплазма базофильна, ядро бледное. Хромосомы деконденсированы, на петлях хромосом типа «ламповых щеток» синтезируется ирформационная РНК. Базофилия цитоплазмы обусловлена накоплением всех видов РНК, увеличением количества рибосом.
В стадии большого роста ооцит резко увеличивается в размерах за счет отложения в его цитоплазме желтка, жира и гликогена. Цитоплазма постепенно приобретает оксифилию и окрашивается эозином в розовый цвет.
Самые крупные - зрелые яйцеклетки, мезолецитальные, умеренно телолецитальные, их цитоплазма оксифильна вследствие накопления значительного количества трофического материала в период большого роста.
Во многих зрелых яйцеклетках в крупном бледно окрашенном ядре с фестончатыми очертаниями обнаруживается большое количество копий рибосомных генов в виде ядрышек, расположенных, преимущественно, на периферии под ядерной оболочкой, накопленных в стадии малого роста в результате амплификации генома. Структурная организация ядра свидетельствует о генетической активности наследственного материала. На многих срезах около ядра видны неокрашенные щели различной величины. Это артефакт, возникающий при сжатии цитоплазмы во время приготовления препарата.
На всех этапах стадии роста яйцеклетки окружены слоем фолликулярных клеток с интенсивно окрашенными гематоксилином в синий цвет ядрами округлой или овальной формы.Вокруг наиболее зрелых яйцеклеток плоские фолликулярные клетки образуют один - два слоя; у яйцеклеток средней величины форма фолликулярных клеток приближается к кубической, и они расположены в два-три слоя; у самых мелких половых клеток фолликулярные клетки уплотнены и формируют лишь один слой.

Рис. 4. Мезолецитальные, умеренно-телолецитальные яйцеклетки амфибии.
На малом увеличении необходимо найти яйцеклетки разной степени зрелости, ориентируясь на цвет цитоплазмы, при большом увеличении следует рассмотреть строение зрелых яйцеклеток.
Препарат № 3. «Яйцеклетка млекопитающего. Яичник кошки» (рис. 5).
Гистологический срез, окраска гематоксилином и эозином.
На срезе яичника кошки под однослойным эпителием и белочной оболочкой в корковом веществе видны ярко окрашенные фолликулы, размеры и организация которых зависят от степени их зрелости. Здесь же присутствуют атретические тела – результат гибели в процессе роста и развития многих фолликулов, в которых можно наблюдать разрушенную яйцеклетку и сморщенную блестящую оболочку под слоем видоизмененных интерстициальных клеток. Клетки зернистого слоя в таких структурах не выявляются вследствии их атрофии. Встречаются белые тела – соединительно-тканные структуры, возникающие как результат преобразования граафова пузырька после овуляции яйцеклетки.
В яичнике млекопитающего яйцеклетки находятся на различных стадиях развития. Размножение женских половых клеток происходит в половой железе зародыша. В постнатальный период, главным образом после полового созревания, в яичнике яйцеклетки находятся в состоянии роста В наружных отделах яичника - под поверхностным эпителием и белочной оболочкой - располагаются примордиалъные фолликулы, содержащие мелкие яйцеклетки на ранних стадиях развития - оогонии, окруженные одним слоем плоских фолликулярных клеток. Вблизи примордиальных находятся растущие фолликулы (первичные), в них находятся ооциты первого порядка. По мере роста фолликулов увеличиваются размеры ооцитов, изменяется фолликулярное окружение. Первоначально уплощенные фолликулярные клетки становятся кубическими и цилиндрическими, из них позже формируются многие слои (вторичные фолликулы). Далее между фолликулярными клетками появляются полости, заполненные серозной жидкостью. Многочисленные полости сливаются между собой с формированием крупного пузырька, который называется граафовым пузырьком (третичным, или преовуляционным фолликулом). В этих образованиях яйцеклетки находятся в яйценосных бугорках, выступающих в полость фолликула.

Рис. 5. Алецитальная, вторично-изолецитальная яйцеклетка млекопитающего.
Строение яйцеклеток млекопитающих (алецитальные, вторично- изолецитальные) наиболее целесообразно изучать в растущих фолликулах или граафовых пузырьках. У них светлое пузыревидное ядро с нежной сеточкой гетерохроматина и ядрышко, цитоплазма окрашена бледно. За цитоплазматической мембраной находится блестящая оболочка, окраска которой может варьировать - от интенсивно-розовой до очень бледной. Яйцеклетки окружены несколькими слоями фолликулярных клеток, образующими вокруг них лучистый венец. В полостях граафовых пузырьков находится жидкость, содержащая гормоны (эстрогены). За наружным слоем фолликулярных клеток, ограничивающих весь пузырек, находятся соединительнотканные оболочки.
На малом увеличении следует найти фолликул, внутри которого находится яйцеклетка с окружающей ее блестящей оболочкой. При большом увеличении рассмотреть строение яйцеклетки.
Препарат № 4. «Сперматозоиды морской свинки» (рис. 6).
Гистологический срез, окраска железным гематоксилином по Гайденгайну.
На препарате мазка спермы при большом увеличении можно увидеть значительное количество сперматозоидов, которые лежат поодиночке или образуют плотные скопления. Часто они склеиваются головками, однако их хвостики разделены, отчего складывается впечатление многохвостности сперматозоидов.
Надо найти участок с отдельно лежащими клетками. Вследствие малых размеров строение сперматозоидов следует изучать при больших увеличениях, лучше - с использованием иммерсии.
В сперматозоидах различают четыре отдела: головку, шейку, перешеек (связующий отдел) и хвостик.
Головки сперматозоидов морской свинки грушевидной формы, содержат интенсивно окрашенную акросому в виде плотной темноокрашенной шапочки, прилежащей к ядру. Ядра занимают большую часть головки, бедны хрматином и выглядят более светлыми в сравнении с акросомой.
За головкой располагается шейка, в цитоплазме которой располагаются центриоли, имеющие вид очень мелких темных точек.
В следующем за шейкой утолщенном перешейке (иначе - связующем отделе) содержатся осевая нить хвостика, вокруг которой по спирали лежат многочисленные митохондрии. В цитоплазме перешейка имеются гранулы гликогена и другие макроэргические вещества, обеспечивающие сперматозоид энергией.
Главный отдел хвостика сперматозоида состоит из осевой нити и окружающей его цитиплазмы, содержащей фермент аденозинтрифосфатаз, расщепляющую АТФ. В более током концевом отделе хвостика осевая нить покрыта только плазмалеммой.
На большом увеличении следует найти отдельно лежащий сперматозоид, рассмотреть особенности его строения. На рисунке (фотографии) отметить отделы сперматозоида: головку с акросомой и ядром, шейку, перешеек (связующий отдел) с митохондриями, хвостик.

А

Б
Рис. 6. Сперматозоиды морской свинки при малом (А) и большом (Б) увеличениях микроскопа. Микропрепарат. Окр. железным гематоксилином по Гайденгайну.
Препарат № 5. «Синкарион лошадиной аскариды» (рис. 7).
Гистологический срез, окраска железным гематоксилином.
На препарате представлен поперечный срез матки лошадиной аскариды, которая выстлана многослойным эпителием. В просвете матки находятся многочисленные оплодотворенные яйцеклетки, окруженные толстыми оболочками оплодотворения, под которыми находится перивителлиновое (околожелточное) пространство и редуционные тельца, если они попали в срез.
После завершения второго редукционного деления гаплоидные родительские геномы одеваются ядерными оболочками — формируются мужской и женский пронуклеусы, которые начинают сближаться, двигаясь по цитоплазме зиготы по сложной траектории. На большом увеличении в центре яйцеклеток, только что завершивших созревание, мелкозернистая ячеистая цитоплазма содержит два пронуклеуса, мужской и женский, до их слияния между собой, каждый из которых включает гаплоидный набор хромосом. Пронуклеусы выглядят как два клеточных ядра. Из-за этого в старых учебниках зиготу на стадии пронуклеусов часто называли термином «синкарион». Различить мужской пронуклеус от женского не удается. Оба пронуклеуса напоминают по строению интерфазные ядра, в которых видны оболочка, хромосомные нити, глыбки хроматина и ядрышки. В одних яйцеклетках пронуклеусы соприкасаются, в других удалены друг от друга. У некоторых пронуклеусов оболочка деформирована.
У некоторых зигот можно видеть остатки двух редукционных телец, причем первое лежит на внутренней стороне оболочки оплодотворения, а второе — на плазматической мембране зиготы. На этой стадии отчетливо видна оболочка оплодотворения, покрытая толстой кутикулой.
В некоторых яйцах можно видеть объединение хромосом в общую метафазную фигуру, что свидетельствует о завершении стадии оплодотворения и переходе к первому митотическому делению, которым начинается стадия дробления. На срезе встречаются также структуры, в которых под оболочку оплодотворения заключены две отдельные клетки - бластомеры. Не следует путать стадию оплодотворения (синкариона) и стадию двух бластомеров.

Рис. 7. Яйцеклетки лошадиной аскариды после оплодотворения. Стадия синкариона. Мужской и женский пронуклеусы в цитоплазме яйцеклетки. Микропрепарат. Окр. окраска железным гематоксилином.
На малом увеличении следует найти структуру, соответствующую стадии синкариона. При большом увеличении необходимо рассмотреть строение синкариона, отметить оболочку оплодотворения, перивителлиновое пространство, редукционные тельца, два пронуклеуса, лежащие в цитоплазме оплодотворенной яйцеклетки.
Тема «Дробление»
Вопросы для самоконтроля:
1. Общие особенности дробления. Основные характеристики стадии дробления.
2. Пространственные типы дробления. Их особенности.
3. Характеристика синхронного периода дробления.
4. Характеристика асинхронного периода дробления.
5. Особенности дробления зигот различных представителей животных в зависимости от количества и распределения желтка.
6. Типы бластул, их строение и особенности формирования у отдельных представителей животного мира.
Препарат № 1. Дробление яйцеклетки лошадиной аскариды (рис. 8).
Гистологический срез, окраска железным гематоксилином по Гайденгайну.
На поперечном срезе матки лошадиной аскариды видны яйцеклетки, находящиеся на различных стадиях дробления. Встречаются также структуры, которые являются синкарионами (строение синкариона представлено на рис. 7).
Сперматозоид вносит в яйцеклетку свое ядро, центросому, митохондрии. Центросома делится, между центриолями образуется веретено, вокруг которого появляется лучистая сфера. Оболочки обоих пронуклеусов при соприкосновении растворяются и хромосомы объединяются в единую группу. Так как каждое ядро привносит по гаплоидному набору хромосом, после объединения восстанавливается диплоидное число хромосом, свойственное всем соматическим и незрелым половым клеткам.
Стадии митоза во время периода дробления имеют ряд морфологических особенностей. В метафазе митотический аппарат деления уже четко виден. Он состоит из веретена и лучистого сияния, которые образованы микротрубочками, отходящими от центриолей. Хромосомы располагаются по экватору веретена. Каждая хромосома состоит из двух хроматид (сестринских хромосом), образование которых произошло путем редупликации в синтетическом периоде интерфазы. У лошадиной аскариды четыре хромосомы.
В анафазе сестринские хроматиды расходятся к противоположным полюсам клетки. В конце анафазы по периферии клетки (в ее центральной части) появляется борозда, которая постепенно углубляется и в телофазе делит тело клетки на две части. Одновременно в телофазе идет реконструкция дочерних ядер. При этом хромосомы деконденсируются, образуются ядрышко и ядерная оболочка.
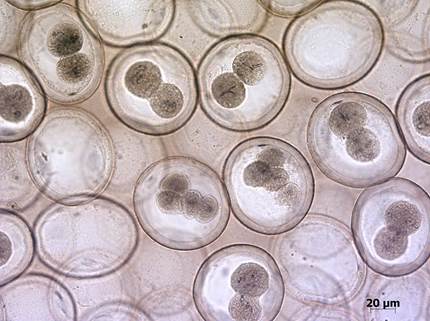
Рис. 8. Дробление яйцеклетки лошадиной аскариды. Различимы стадии 2-х и 4-х бластомеров. Микропрепарат. Окр. железным гематоксилином.
Стадия образования двух бластомеров. Среди зигот на стадиях анафазы и телофазы первого деления дробления располагаются отдельные эмбрионы на стадии двух бластомеров, уже завершившие первое деление. Эту стадию нужно уметь отличать от поздней телофазы. На двуклеточной стадии бластомеры полностью отделены друг от друга и часто лежат в разных оптических плоскостях, а на стадии телофазы обычно хорошо видны борозда дробления и контрактильное кольцо.
Бластомеры одинаковой величины, что указывает на полное и равномерное дробление. В ячеистой цитоплазме бластомеров видно интерфазное ядро с глыбками гетерохроматина.
У лошадиной аскариды билатеральный тип дробления, который приводит к образованию на стадии четырех бластомеров вначале Т-образной, а затем ромбовидной структуры вследствие пространственной переориентации бластомеров.
На малом и большом увеличениях следует найти яйцеклетку на стадии 2-х, 4-х или большего количества бластомеров. Обратить внимание на наличие оболочки оплодотворения формирующейся в перивителлиновом пространстве бластулы, а также присутствующие там редукционные тельца.
Препарат № 2. Дробление умеренно телолецитальной яйцеклетки лягушки (рис. 9).
Гистологический срез, окраска по Ван-Гизону.
Яйцеклетки амфибий содержат среднее количество желтка, который неравномерно распределен вдоль анимально-вегетативной оси яйца (мезолецитальные, умеренно телолецитальные яйцеклетки), претерпевают полное (голобластическое) неравномерное, радиальное дробление. В результате такого дробления возникает зародыш — амфибластула.
Для мезолецитальных яйцеклеток ортогональность борозд первых трех делений дробления выводится непосредственно из правил Гертвига - Сакса. Веретено первого деления дробления в зиготе амфибий располагается широтно под поверхностью анимального полюса. Соответственно борозда первого деления дробления проходит меридионально и называется меридиональной. Меридиональная борозда закладывается на анимальном полюсе и медленно распространяется на вегетативную область. Она делит серый серп, образовавшийся в результате ооплазматической сегрегации. Желток, сосредоточенный в вегетативном полушарии, затрудняет дробление.
Борозда второго деления начинает закладываться вблизи от анимального полюса еще до того, как борозда первого деления дробления разделила вегетативное полушарие. Веретена второго деления расположены в той же плоскости, что и веретено первого деления, но под прямым углом к нему, поскольку эти направления примерно соответствуют наибольшему протяжению свободной от желтка цитоплазмы. Борозда второго деления дробления также меридиональная.
Первые четыре бластомера мезолецитальных яйцеклеток примерно равны между собой. Далее направление наибольшей протяженности свободной цитоплазмы в каждом бластомере совпадает с меридианами яйца, так как широтные поперечники короче меридиональных. Соответственно этому все четыре веретена третьего деления располагаются меридионально, но остаются смещенными к анимальному полюсу. Вследствие этого б орозды третьего деления дробления проходят широтно и смещены к анимальному полюсу. В результате образуются четыре анимальных бластомера меньшего размера и четыре более крупных вегетативных бластомера.
Далее чередуются меридиональные и широтные борозды, а также появляется еще одно направление дробления – тангенциальное, которое происходит внутри бластулы параллельно поверхности зародыша.
Позже дробление утрачивает общую правильность, и на последующих стадиях дробления анимальная область содержит многочисленные мелкие клетки, а в вегетативной области имеется относительно небольшое количество крупных, богатых желтком бластомеров.
Иногда между немногочисленными бластомерами имеется небольшая полость – полость Бэра.
Поскольку субкортикальная зона анимального полушария яйцеклеток амфибий и осетровых рыб содержит многочисленные пигментные гранулы, анимальные бластомеры дробящихся зародышей имеют более темную окраску. Клетки-потомки пигментированных анимальных бластомеров сохраняют окраску в течение всего раннего развития, вплоть до стадии нейрулы.

Рис. 9. Дробление умеренно-телолецитальной яйцеклетки амфибии. Микропрепарат. Окр. по Ван-Гизону.
На малом увеличении необходимо определить типы имеющихся борозд дробления. При этом следует учитывать степень пигментации бластомеров и их величину.
Препарат № 3. Амфибластула лягушки. Меридиональный срез (рис. 10).
Гистологический срез, окраска по Ван-Гизону.
На данном препарате под микроскопом при малом увеличении представлен общий вид амфибластулы на меридиональном срезе.
Образующаяся амфибластула характеризуется следующими признаками:
1) форма сферическая;
2) стенка состоит из нескольких слоев клеток;
3) бластоцель смещен к анимальному полюсу;
4) анимальная часть стенки бластулы (крыша бластоцеля) тоньше, чем массивная вегетативная часть стенки (дно бластоцеля);
5) анимальные бластомеры пигментированы, содержат меньшее количество желтка, чем крупные вегетативные бластомеры, и мельче их.
Анимальная часть стенки амфибластулы (крыша бластоцеля) тоньше, чем массивная вегетативная часть стенки (дно бластоцеля). Между ними по бокам от бластоцеля расположена краевая зона. Более мелкие анимальные бластомеры (микромеры) сильнее пигментированы, содержат меньшее количество желтка и мельче, чем крупные вегетативные бластомеры (макромеры). Бластомеры краевой зоны промежуточного размера (мезомеры).
Микромеры, относящиеся к разным слоям, составляющим крышу бластоцеля, пигментированы по-разному. Для микромеров, кроме приядерного расположения пигмента, характерно также наличие слоя пигмента вдоль внешнего края клеток. Этот пигмент кортикальной области анимальной части ооцита унаследовали клетки, которые при дроблении сформировались из этой зиготы. В ранней амфибластуле, когда крыша бластоцеля состоит в основном из одного слоя клеток, слой пигмента есть во всех микромерах. В поздней амфибластуле, когда крыша бластоцеля становится многослойной, пигмент наследуют только клетки внешнего слоя. При дальнейшем развитии (стадии гаструляции и нейруляции) можно проследить судьбу потомков этих клеток по наличию пигмента в кортикальной области.
При изучении препарата амфибластулы рекомендуется расположить ее анимальным полюсом вверх. Следует рассмотреть особенности бластомеров различных частей бластулы. На большом увеличении нужно внимательно рассмотреть крышу бластоцеля, обращая внимание на распределение пигмента в клетках. Отметить расположение крыши, дна и краевой зоны бластулы, а также бластоцеля, слегка смещенного к анимальному полюсу.

Рис. 10. Амфибластула лягушки. Микропрепарат. Окр. по Ван-Гизону.
Тема «Гаструляция у зародышей беззубки и амфибии.
Нейруляция»
Вопросы для самоконтроля:
1. Понятие гаструляции. Биологическое значение гаструляции.
2. Основные способы гаструляции.
3. Закладка мезодермы у низших позвоночных. Дифференцировка мезодермы.
4. Гаструляция у беззубки.
5. Гаструляция у амфибии.
6. Механизм нейруляции.
7. Карта презумптивных участков зародыша амфибии.
Препараты № 1-3. Гаструляция у зародыша лягушки. Саггитальный срез (рис. 11, 12, 13).
Гистологические срезы, окраска по Ван-Гизону.
На препаратах 1-3 представлены саггитальные срезы ранней, средней и поздней гаструлы лягушки. Их изучение позволит составить представление о последовательности событий и механизме процесса формирования трех зародышевых листков во время гаструляции у амфибий.
На срезе ранней гаструлы (рис. 11) отчетливо видна зона, где произошла попытка к инвагинации, с которой начинается гаструляция у амфибий. Это область серого серпа. В результате наметилась дорзальная губа бластопора, клетки которой интенсивно пигментированы. В области образованной зарубки имеются колбовидные клетки, которые хорошо различимы благодаря характерной форме. Клетки погружаются вглубь, сохраняя связь с поверхностью.
Поверхностные клетки анимального полюса содержат гранулы пигмента под наружной мембраной. В крупных клетках вегетативного полюса пигмент отсутствует. Щелевидный бластопор срезан поперек. Его закругленный анимальный край и есть дорсальная губа бластопора, через которую подворачивается материал краевой зоны, который в дальнейшем образует прехордальную пластинку (материал передней кишки), хорду и головную мезодерму.
Клетки поверхностного и подповерхностных слоев сначала смещаются вегетативно, а затем вместе с колбовидными клетками распластываются и ползут в анимальном направлении по внутренней стенке крыши бластоцеля. Этот процесс называется инволюцией. С вентральной стороны зародыша подобных движений пока нет.
Одновременно с этим происходит активная пролиферация бластомеров анимального полюса, которые начинают движение в вегето-петальном направлении (от анимального полюса к вегетативному), обрастая мезо- и макромеры снаружи. Этот процесс называется эпиболией. Эпиболия приводит к образованию энтодермы и эктодермы.

Рис. 11. Гаструляция лягушки. Ранняя гаструла. Образование дорсальной губы бластопора. Микропрепарат. Окр. по Ван-Гизону.
На стадии средней гаструлы (рис. 12) на дорсальной стороне зародыша клетки краевой зоны продолжают подворачиваться внутрь зародыша. Образующийся гастроцель (полость первичной кишки, архентерон) становится глубже. Его дорсальная стенка (или крыша) образована подвернувшимися клетками поверхностного слоя краевой зоны, а вентральная стенка (или дно) — погрузившимися клетками эпиболирующей массы, увлекающей за собой непигментированные бластомеры с желтком вегетативного полюса. Одновременно с описанными выше процессами происходит подворачивание боковых бластомеров, которые в дальнейшем дают мезодерму, с образованием латеральных (боковых) губ бластопора. Часть бластомеров с желтком некоторое время заполняют бластопор, образуя желточную пробку, некоторое время выступающую наружу.
Бластоцель в результате миграции клеток через губы бластопора уменьшается в размерах, принимает вид узкой щели и начинает смещаться образующимся гастроцелем в вегетативную область гаструлы.

Рис. 12. Гаструляция лягушки. Средняя гаструла. Формирование желточной пробки между губами бластопора. Микропрепарат. Окр. по Ван-Гизону.
На стадии поздней гаструлы (рис. 13) лидирующий край дорсальной мезодермы достигает анимального полюса. В результате латеро-медиальной миграции клеток во всех слоях краевой зоны в дорсальном направлении и их конвергентного встраивания спинная сторона зародыша вытягивается. Колбовидные клетки образуют эпителиальную выстилку архентерона в головной части. Бластоцель существенно уменьшается в размерах и полностью смещается на вентральную сторону зародыша. Часто на препаратах он не обнаруживается. Вентральная губа становится незначительно глубже. Бластопор сокращается в диаметре благодаря эпиболии, конвергентному растяжению и погружению клеток желточной пробки. Сокращается количество слоев клеток в мезодерме. На дорсальной стороне в подвернувшейся мезодерме можно видеть начало формирования хорды.

Рис. 13. Гаструляция лягушки. Поздняя гаструла. Исчезновение желточной пробки между губами бластопора, формирование гастроцеля. Микропрепарат. Окр. по Ван-Гизону.
Рассмотрите при малом увеличении сагиттальные срезы ранней, средней и поздней гаструлы.
Определите на препарате анимальный и вегетативный полюса зародыша, а также его дорсальную и вентральную губы бластопора. Сравните смещение в результате эпиболии анимального пигментированного материала на дорсальной и вентральной сторонах.
Обратите внимание на изменение положения бластоцеля. Найдите полости гастроцеля, бластоцеля, желточную пробку.
Препараты № 4-6. Нейруляция у зародыша лягушки. Поперечный срез (рис. 14, 15, 16).
Гистологические срезы, окраска по Ван-Гизону
Date: 2015-10-19; view: 15103; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |