Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Электрохимического потенциала
Важнейшим интермедиатом в реакциях превращения энергии в биологических системах является трансмембранный градиент электрохимического потенциала. В соответствии с хемиосмотической теорией П. Митчелла и развиваемой в настоящее время ротационной теорией П. Бойера АТФ синтезируется в хлоропластах в результате сложного комплекса сопряженных процессов: транспорта электронов, генерации электрохимического потенциала и преобразования его энергии в серии конформационных изменений отдельных субъединиц сопрягающего комплекса, передачи конформационных сигналов в каталитические центры.
Механизм сопряжения электронного транспорта с генерацией электрохимического потенциала рассматривается в настоящее время в рамках хемиосмотической теории Митчелла (теории протонного сопряжения), сформулированной в 1961 г. и развитой в последующие годы (1966). Теория Митчелла включает два основных положения: 1) энергия, освобождаемая в редокс-реакциях, первично запасается в форме протонного градиента (∆рН); 2) признается необходимость сопрягающей мембраны, непроницаемой для Н+. При работе ЭТЦ сопряженно с транспортом электронов происходит энергозависимый перенос протонов через гидрофобный слой мембраны из стромы в люмен, направленный против электрического и концентрационного градиента.
Сопрягающей мембране в теории Митчелла отводится ведущая роль в процессе фотофосфорилирования. Благодаря тому, что мембрана непроницаема для протонов, может быть создана разность свободных энергий в двух водных фазах, разделенных мембраной. Тилакоидная мембрана ориентирует комплексы, участвующие в транспорте электронов, таким образом, что создается векторный поток электронов и протонов через мембрану. В результате сопряженных процессов переноса электронов (на стромальную поверхность мембраны) и Н+ (внутрь тилакоидов) возникает градиент протонов (∆рН), который служит источником энергии для синтеза АТФ. Значительный вклад в суммарное значение разности энергий двух фаз, разделенных мембраной (строма, люмен), дает образующийся при работе ЭТЦ электрический мембранный потенциал. Электрическое поле на мембране образуется в течение 10 мс при первичном фотохимическом разделении зарядов.
В результате определенного расположения и взаимодействия участников редокс-реакций в мембране редокс-энергия непосредственно преобразуется в энергию электрохимического потенциала (Δ 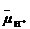 ), который представляет собой сумму электрической (∆ψ) и осмотической (∆рН) энергии и является источником свободной энергии для клетки:
), который представляет собой сумму электрической (∆ψ) и осмотической (∆рН) энергии и является источником свободной энергии для клетки:
Δ = -2,3lgT∆pH + F ∆ψ.
Электрохимический трансмембранный градиент Δ формируется в результате сопряжения электрогенных и протолитических реакций при работе ЭТЦ. В нециклическом фотофосфорилировании известны три электрогенные реакции, в результате которых происходит перенос электронов с внутренней на наружную, стромальную поверхность мембраны с запасанием энергии в форме электрического мембранного потенциала. Две реакции имеют место в реакционных центрах ФС1 и ФСII. Они связаны с быстрым переносом электрона от фотовозбужденного пигмента к конечному компоненту акцепторного комплекса, локализованному на стромальной поверхности мембраны. Одна, медленная электрогенная реакция связана с работой Q-цикла, с переносом электрона от молекулы PQH2, окисляемой Fe-S-белком Риске, к высокопотенциальному цитохрому b6.
При работе ЭТЦ электрогенные реакции сопряжены с протолитическими. В реакциях трансмембранного переноса протонов из стромального пространства в люменальное значительная роль принадлежит цитохромному b6 f -комплексу. При окислении пластохинола в центре Qz цитохромного комплекса удаление двух электронов из молекулы PQH2 сопряжено с освобождением двух Н+, которые поступают в люмен, а окисленный пластохинон может быть снова восстановлен в QB центре (ФСII) или QC-центре (цитохромный b6 f -комплекс) на стромальной стороне мембраны с использованием двух протонов из стромы. Другая реакция образования протонного градиента обусловлена процессом фотоокисления воды ФСII, в результате которого при разложении двух молекул Н2О выделяется молекулярный кислород, электроны поступают в ЭТЦ, а в люменальное пространство высвобождаются четыре протона. Перенос протонов через мембрану происходит также при циклическом транспорте электронов вокруг ФСII при участии цитохрома b559.
Таким образом, энергия, освобождаемая при переносе электронов в отдельных участках ЭТЦ, преобразуется в осмотическую и электрическую энергию градиента электрохимического потенциала.
Градиент электрохимического потенциала, согласно ротационной теории П.Бойера (1993, 1997), служит источником энергии для конформационных изменений белков АТФ-синтазного комплекса CF0-CF1 в результате которых в каталитическом центре сопрягающего фактора I (CF1 образуется молекула АТФиз АДФ и ФH.
Date: 2015-07-22; view: 707; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |