Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Поведенческие реакции и секреция АКТГ 5 page
Эти наблюдения свидетельствуют о существовании различных форм трофотропного состояния, характерных
ТОШНОТА И ЛИХОРАДОЧНОЕ СОСТОЯНИЕ
для сна, тошноты и сосудистого коллапса. Но при любом таком состоянии тонус симпатического отдела гипоталамуса понижен, а парасимпатического — повышен. Уменьшение гипоталамо-кортикальных влияний и мышечного тонуса обусловлено снижением эрготропных функций заднего отдела гипоталамуса. Высвобождающаяся парасимпатическая активность переднего отдела гипоталамуса совместно со снижением симпатических разрядов вызывает в этих условиях падение кровяного давления, замедление сердечного ритма и уменьшение минутного объема. Вследствие снижения гипоталамо-кортикальных влияний в ЭЭГ при сне и коллапсе возникают медленные потенциалы; при тошноте уменьшается сенсорная и эмоциональная реактивность, а после обильной еды затормаживается умственная деятельность.
Вопрос о связи эмоций с тошнотой требует дальнейших пояснений. Уже упоминалось, что состояние страха нередко сопровождается тошнотой и рвотой, но несомненно, что обычной реакцией па страх является попытка избежать опасности. Эта попытка оказывается несостоятельной, если страх сопровождается тошнотой. Головокружение и снижение мышечного тонуса приводит к ослаблению зрительного восприятия и снижению скорости, устойчивости и координации двигательных реакций. При наличии реальной возможности избежать опасности страх обычно не сопровождается тошнотой. Если же по тем или иным причинам избежать опасности нельзя, возникает тошнота, свидетельствуя о высокой интенсивности страха и (или) о наличии опасности для жизни. Это относится не только к человеку, но и к экспериментальным животным в условиях конфликтной ситуации, связанной с возбуждением. Синдром тошноты и рвоты с сосудистым коллапсом может возникать также в тех случаях, когда человеку угрожает опасность психологического или социального характера [716]. Хотя мы не можем проследить, как распространяется в мозге возбуждение, сигнализирующее нам о критической ситуации, но ясно одно, что лежащее в его основе эмоциональное возбуждение включает кортикофугальные импульсы к гипоталамусу. Мы не располагаем данными о характере коркового возбуждения и о том, какие области коры участвуют в осу-
ГЛАВА XVI
ществлении этих процессов, по можно предположить, что к таким областям относится передняя часть поясной извилины, так как раздражение этой структуры у обезьян вызывает, кроме возбуждения вегетативной системы, полное расслабление мускулатуры [848], а ее удаление устраняет, по крайней мере временно, реакцию страха [521, 929]. Нам бы хотелось подчеркнуть тонкую приспособляемость организма к разным эмоциональным ситуациям; так, вегетативные, соматические и поведенческие реакции при страхе совершенно различны в зависимости от того, реальна ли возможность избежать опаспости или нет.
ПРИСПОСОБИТЕЛЬНЫЕ РЕАКЦИИ ПРИ ЛИХОРАДОЧНЫХ
состояниях
Тошнота относится к состояниям, лежащим на грани нормы и патологии. Интересно поэтому кратко проанализировать такое общее патологическое состояние, пр' котором нарушается ответ всего организма в целом, что бы определить, применима ли точка зрения, основанна на предшествующем анализе физиологических исследований, к этой ситуации. В качестве примера нами был выбрано лихорадочное состояние — не только в силу ег большого значения, но также и потому, что в его возникновении участвует гипоталамус, который связан с регуляцией температуры тела.
Со времени классических исследований Рансон [735] и его сотрудников известно, что задний отдел гипо таламуса регулирует теплообразование, а передний от дел — теплоотдачу посредством расширения сосудов одышки.
Изучение лихорадочного состояния в клинике пока зало, что оно обусловлено снижением теплоотдачи. Эт подтверждено экспериментальными исследованиями, которых производили двухсторонние разрушения в об ласти между перекрестом зрительных нервов и передней комиссурои. При распространении этих разрушений в каудальном направлении на передний отдел гипоталамуса отмечалось быстрое повышение температуры до 41° и смерть [49а]. При неполном разрушении переднего отдела
ТОШНОТА И ЛИХОРАДОЧНОЕ СОСТОЯНИЕ
гипоталамуса температура не достигала столь высоких значений. О нарушении механизма теплоотдачи у таких животных свидетельствует факт значительного повышения температуры тела при повышении температуры окружающего воздуха. Напротив, терморегуляция в условиях холода, осуществляемая задним отделом гипоталамуса, оставалась нормальной. Клинические наблюдения подтверждают, что разрушение переднего отдела гипоталамуса у человека приводит к резкому повышению температуры с летальным исходом [49].
При обширных разрушениях переднего отдела гипоталамуса возникает лихорадочное состояние с ускорением частоты сердечных сокращений и пилоэрекцией, что свидетельствует о высвобождении симпатических механизмов заднего отдела гипоталамуса. Более того, в этих экспериментах наблюдалась дрожь, возникающая, как известно, при раздражении заднего отдела гипоталамуса [66] и исчезающая при его удалении [735]. Отсюда явствует, что при экспериментально вызванном лихорадочном состоянии происходит высвобождение заднего отдела гипоталамуса, распространяющееся и на соматическую систему, о чем свидетельствует появление дрожи.
Фармакологические исследования подтверждают, что в основе повышения температуры лежит повышение активности симпатических центров. Центральные стимуляторы симпатической системы вызывают лихорадочное состояние либо за счет усиленной теплопродукции, либо за счет пониженной теплоотдачи [849].
Как отмечалось выше, задний отдел гипоталамуса и ретикулярная формация отличаются высокой чувствительностью к подавляющему действию барбитуратов [264, 304]. Этим можно объяснить временное снятие лихорадочного состояния, возникшего при разрушении переднего отдела гипоталамуса внутривенной инъекцией барбитуратов [49а]. Это еще раз указывает на роль эр-готропных систем (заднего отдела гипоталамуса и активирующей ретикулярной формации) в геиезе лихорадочного состояния и подтверждает положение о том, что их активация при разрушении переднего отдела гипоталамуса в значительной степени ответственна за последующее повышение температуры. Температура тела за-
ГЛАВА XVI
висит от равновесия процессов теплообразования и удержания тепла, с одной стороны, и теплоотдачи — с другой. Эти противоположно направленные процессы в основном регулируются соответственно задним и передним отделами гипоталамуса. Если активность этих отделов находится в равновесии, то температура тела удерживается в пределах нормы. Лихорадочное состояние свидетельствует о патологическом преобладании симпатической системы; такое преобладание может быть следствием либо повышения активности симпатической системы, либо понижения активности парасимпатической системы, либо, наконец, следствием обоих процессов. Разрушение переднего отдела гипоталамуса приводит к нарушению теплоотдачи и, кроме того, к высвобождению заднего отдела гипоталамуса,активация которого усиливает теплообразование и способствует удержанию тепла. Поэтому температура тела повышается. Преобладание симпатической системы, наблюдаемое в этих условиях, оказывается даже более выраженным, чем можно было бы ожидать на основании одного лишь «высвобождения», поскольку реактивность симпатических центров гипоталамуса при повышении температуры возрастает. Сходные сдвиги гипоталамического равновесия в сторону резкого преобладания симпатической системы лежат, вероятно, и в основе генеза лихорадочного состояния, возникающего под влиянием некоторых фармакологических веществ или при различных заболеваниях. Подобное нарушение равновесия может влиять не только на температуру тела, но и на поведение организма в целом. I Теперь на основании упомянутых физиологических исследований попытаемся истолковать клинические данные, касающиеся лихорадочного состояния. Клинический опыт слагается из наблюдений над больными с лихорадочным состоянием при инфекционных заболеваниях, а также из исследований с введением пирогепных веществ (вызывающих лихорадку). В последнем случае, в зависимости от дозы препарата можно вызвать различную степень повышения температуры. При сопоставлении данных ряда авторов создается впечатление, чт при повышении температуры па долю градуса эмоциональная реактивность возрастает [508, 707, 801]. Неболь-
ТОШНОТА И ЛИХОРАДОЧНОЕ СОСТОЯНИЕ 385
шое повышение температуры сразу же сказывается на настроении больного — минимальный раздражитель вызывает недовольство и даже приводит в ярость. Лишь пои значительном повышении температуры возникает состояние летаргии и затемнение сознания. Эти наблюдения указывают на активацию заднего отдела гипоталамуса при умеренном повышении температуры, хотя при высокой температуре реактивность мозга снижается.
Часто наблюдаемая при лихорадочном состоянии повышенная реактивность симпатического отдела гипоталамуса, сопровождающаяся активацией соматической нервной системы, иллюстрируется двигательным беспокойством и повышением реактивности к сенсорным раздражителям. Раздражающее действие света и звука, даже при малой их интенсивности, объясняется, видимо, повышенной эмоциональной реактивностью, а не возросшей чувствительностью сенсорных механизмов.
Икскюль указывал на состояние покоя как на первостепенную необходимость для организма при лихорадке; однако эта цель достигается не пассивным поведением, а активными попытками восстановить состояние психического и физического покоя. Подобная активность характеризуется не только возросшей скоростью, но и меньшей правильностью при выполнении психологических тестов. При тошноте, напротив, преобладает состояние пассивности, как бы предохраняющее организм от воздействия внешних раздражителей. Очень интересно, что при этих чисто интуитивных объяснениях поведения человека при тошноте и лихорадке фигурируют указания на различия, обусловленные разным состоянием эрго-тропиой системы при этих двух патологических проявлениях.
Лихорадочное состояние сопровождается нарушением психических функций. Замедляются ассоциативные процессы; изменения почерка свидетельствуют о проприо-нептивных нарушениях. Все это указывает на ослабление нервных функций и связано, видимо, с уменьшением частоты и амплитуды корковых потенциалов при повышенной температуре [548]. Может иметь значение и некоторая степень аноксии коры. При повышении температуры повышается интенсивность обмена в тканях
13—517
ГЛАВА XVI
мозга, а кора более чувствительна к недостатку кислорода, чем подкорковые структуры. Роль относительной аноксии лучше всего демонстрируется при сравнении влияния повышения температуры на кору мозга в норме и при судорожной активности в ней. Эти опыты показывают, что судорожные разряды уменьшаются по амплитуде и даже исчезают при той температуре, при которой нормальные потенциалы коры не изменяются [889].
При судорожной активности нейроны необычайно чувствительны к аноксии и могут поэтому служить индикатором даже незначительных сдвигов в содержании кислорода, значение которых для осуществления сложных корковых процессов хорошо известно [343, 350].
Таким образом, при лихорадочном состоянии имеет место симпатическая «настройка» гипоталамуса, которая создает оптимальные условия для поддержания мозгового кровообращения, как бы защищая мозг от действия аноксии. Вместе с тем относительная аноксия коры как бы ограничивает рост сенсорной реактивности, под влиянием усиления гипоталамо-кортикальных разрядов. Благодаря этому достигается компромисс — осуществляется защита от мозговых расстройств и чрезмерной реактивности, особенно в отношении боли, которая, как свидетельствуют результаты фронтальной лоботомии, в большой степени зависит от коры мозга. Опыты на животных с локальным действием тепла на кору выявляют большой «запас прочности». Так, нагревание коры мозга кошки до 46° не сопровождается выраженным изменением порога возбудимости моторной коры [888]; соответствующими данными о мозге человека мы не располагаем. Кроме того, следует иметь в виду, что лихорадочное состояние — это не просто повышение температуры.
Мы считаем, что существует широкий диапазон уровней настройки — от физиологических сдвигов, вызываемых рефлексами с барорецепторов, до патологического лихорадочного состояния; тошнота занимает в этом отношении промежуточное положение. Основные сдвиги равновесия систем гипоталамуса, наблюдаемые в этих условиях, испытывают влияние со стороны висцеральных рефлексов, примерами которых могут служить различные трофотропные состояния, возникающие при тошноте,
IHOTA И ЛИХОРАДОЧНОЕ СОСТОЯНИЕ 387
сытости и сосудистом коллапсе. Эрготропное состояние, физиологически проявляющееся при эмоциональном возбуждении (ярость), изменяется при лихорадке в результате действия повышенной температуры на кору, в которой происходят психические процессы. Однако, несмотря на эти изменения, легко выявить основные различия, существующие между разными трофотропными и эрго-тропными состояниями, а также определяющее влияние равновесия систем гипоталамуса на общее поведение организма.
13»
ГЛАВА XVII
ФИЗИОЛОГИЧЕСКИЕ АСПЕКТЫ МЫШЕЧНОГО РАССЛАБЛЕНИЯ
Мы видели, что эмоциональное возбуждение, приводящее к активации гипоталамической системы, лежит в основе многих психосоматических расстройств. В связи с этими наблюдениями возникает вопрос, нельзя ли снизить интенсивность тех физиологических факторов, которые поддерживают возбудимость гипоталамуса, и таким образом избежать патологических последствий.
ФИЗИОЛОГИЧЕСКИЕ ВЛИЯНИЯ СНИЖЕНИЯ МЫШЕЧНОГО НАПРЯЖЕНИЯ
Выше мы упоминали, что реакция пробуждения, выражающаяся в диффузной активации коры мозга и возбуждении гипоталамуса, возникает у кошки, находящейся под легким наркозом, под влиянием проприо- или ноцицептивных раздражений, но не возникает (или возникает лишь в редких случаях) при слуховых, зрительных или тактильных раздражениях. Поскольку в строго физиологических условиях болевые воздействия отсутствуют, то первостепенную роль в поддержании возбудимости гипоталамуса, по-видимому, играют мышечные веретена. Этот вывод проверен в ряде экспериментов [312], в которых мышечные импульсы блокировали с помощью кураре или курареподобных веществ (анектин). При введении этих веществ в малых дозах (не влияющих на кровяное давление) симпатическая реактивность гипоталамуса снижалась. Фиг. 38 иллюстрирует значительно менее выраженное сокращение третьего века и повышение кровяного давления при раздражении заднего отде-
(ЛОГИЧЕСКИЕ АСПЕКТЫ МЫШЕЧНОГО РАССЛАБЛЕНИЯ 389
ла гипоталамуса после введения кураре. Этот эффект отражается на состоянии активности коры головного мозга. У кошки под легким нембуталовым наркозом и при местной анестезии в контрольных условиях возникают редкие группы потенциалов (фиг. 39, А), а после
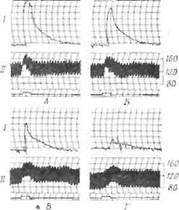
Фиг. 38. Уменьшение симпатической реактивности заднего отдела гипоталамуса при действии интокострина [312].
{„Т т1)етье веко; // — крозяное давление. Характеристика раздражения: 2.5 в. 207 имп/сек. 0.8 мсек, 8 сек. А. Контроль. Б. Через 4 мин после введения интокострина (0,25 мл/кг, внутримышечно). В и Г. Соответственно через 10 и 20 мин после дальнейшего введения 0.1 мл/кг интокострина внутривенно. Отмечается постепенное снижение силы сокращения третьего века и степени гипертонии.
введения анектина регистрируются отчетливые многочисленные группы (как во время сна или при барбиту-ратном наркозе) (фиг. 39, Б). Эти выраженные изменения не сопровождаются сдвигом кровяного давления и полностью обратимы спустя несколько минут после прекращения действия анектина (фиг. 39, В).
ГЛАВА XVII
Интегрированная (с помощью анализатора Оффне-ра) амплитуда потенциалов при действии анектина значительно возрастает в области низких частот. Это отражает синхронизацию нейронных разрядов при снижении возбудимости. Наконец, отмечено, что после введения

Фиг. 39. Влияние анектина на корковые и гипоталамичеекие потенциалы у кошки [312].
Биполярная регистрация: / — левый и правый гипоталамус; 2 — левая моторная и теменная кора; 3 — левая и прабая моторная кора; 4 — частотный анализ гипоталамических потенциалов с помощью анализатора Оффнера. Анализируемые частоты: 1, 5; 2; 2.5; 3; 3.5; 4; 5; 6; 7; 8; 9; 10; 11; 12; 13: 14; 15; 16; 18; 20; 22; 24; 27; 30. А. Контроль. И. Через 3 мин после введения анектина (0,6 мг/кг). П. Через 8. мин после Б.
кураре болевое раздражение вызывает менее выраженное состояние возбуждения в коре и гипоталамусе. Только что описанные три вида экспериментов иллюстрируют разные стороны одного и того же явления. Снижение реактивности гипоталамуса обусловливает менее выраженный симпатический эффект при прямом раздражении заднего отдела гипоталамуса и при рефлекторной активации гипоталамо-кортикалыюй системы болевым раздражением. Поскольку гипоталамус дает тонические разряды, влияющие на степень активации коры в целом, дозы кураре или курареподобных веществ, подавляющие возбудимость гипоталамуса, снижают гипотадамо-
ОЛОГИЧЕСКИЕ АСПЕКТЬ1 МЫШЕЧНОГО РАССЛАБЛЕНИЯ 391
кортикальные разряды, приводя тем самым к появлению групповых потенциалов и сдвигу спектра ЭЭГ в сторону низких частот.
Состояние сниженной возбудимости гипоталамуса бы-iio подтверждено в опытах на ненаркотизированных кошках с вживленными электродами. После введения кураре у кошек наблюдалась сонливость. Реакции пробуждения были укорочены, а проявления ярости при раздражении гипоталамуса снижены, хотя дозы вводимых веществ были слишком малы, чтобы оказывать влияние на дыхание и позу. У животных, которые в контрольных условиях при встрече с лающей собакой проявляли агрессивное поведение, после введения препарата наблюдалась реакция избегания [310]. Такого рода изменения поведения, видимо, также являются выражением сниженной активности гипоталамической системы, поскольку реакции избегания возникают при меньшей интенсивности раздражений, чем агрессивное поведение [491].
Следует добавить, что при расслаблении мышц снижается реактивность к болевым раздражениям, которые, как было показано, активируют гипоталамо-кортикаль-ную систему [129]. Эти факты, видимо, лежат в основе хорошо известной связи, существующей между эмоциональной реактивностью и реактивностью к болевым раздражениям [426].
МЕХАНИЗМ ПОЛОЖИТЕЛЬНОЙ ОБРАТНОЙ СВЯЗИ ПРОПРИОРЕЦЕПТОРОВ
Повышенная активность соматической нервной системы всегда связана как с раздражением заднего отдела гипоталамуса, так и с эмоциональным возбуждением. Выражение лица и поза сильно меняются при различных эмоциональных состояниях. Активность мышечных веретен, возрастающая при этих состояниях (исключение составляет эмоция крайнего ужаса), способна влиять на состояние возбуждения гипоталамуса, вызы-вая дальнейшее усиление его реактивности по отношению к различным раздражителям, а также интенсивности гипоталамо-кортикальных разрядов. Но существуют
m->
ГЛАВА XVII
и другие предпосылки для того, чтобы предположить наличие механизма положительной обратной связи. Из исследований Каффлера и Ханта [556], а также Гранита [393] и других мы знаем, что тонкие нервные волокна иннервируют мышечные веретена и оказывают значительное влияние на их реактивность. При постоянном мышечном напряжении афферентный разряд, возникающий в мышечных веретенах, растет с повышением степени возбуждения в системе тонких волокон. Механизм, с помощью которого это осуществляется, функционирует следующим образом. Система тонких эфферентов иннервирует сократительные участки «интрафузальных» мышечных волокон. Посередине каждого интрафузаль-ного мышечного волокна расположен небольшой несократимый участок, окруженный аннулоспиральными окончаниями, или первичными рецепторами. Это схематически показано на фиг. 40. В ответ на приход импульсов по тонким гамма-волокнам крайние (лежащие у 'полюсов) участки веретена сокращаются. Если мышца как целое не сокращается, конечные участки интрафузальных волокон находятся в относительно фиксированном состоянии, так что центральные области растягиваются и первичные рецепторы генерируют импульсы. Эти афферентные импульсы обеспечивают положительную обратную связь, усиливая активность ретикулярной системы, которая в первую очередь ответственна за возникновение разрядов в гамма-волокнах.
Хотя мы не согласны с «активационной» теорией эмоций Линдсли [580] (см. гл. XXV), не может быть никакого сомнения в том, что эмоциональное возбуждение сопровождается усилением активности ретикулярной формации, существенное влияние на которую оказывают кортикофугальные разряды [5, 260]. Независимо от сложности типов возбуждения, в которых могут участвовать кора, гипоталамус и ретикулярная формация, происходит неизбежное возбуждение системы тонких нервных волокон, которая усиливает разряды, идущие от мышечных веретен и, таким образом, способствует дальнейшему возрастанию гипоталамического (и эмоционального) возбуждения. Этот параллелизм в активности соматической системы и симпатического отдела вегетативной системы
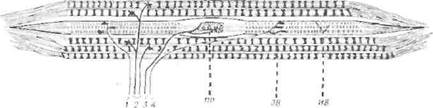
Рис. 40. Схема мышечного веретена, содержащего 3 интрафузальны.х волокна.
Центральный участок этих волокон лишен поперечной исчерчснности и несократим. Окончание первичного рецептора (ПР) обвивает, как спираль, центральную область и генерирует импульсы, идущие по толстым афферентным волокнам. Сократительные краевые участки интрафузальных волокон (ИВ) получают веточки от мелких гамма-мотонейронов (/ и 4); экстрафузальные волокна (ЭВ) иннервируются толстыми волокнами двигательных нервов (2 и 3).
ГЛАВА XVII
наблюдается и при произвольных сокращениях мышц. Так, сгибание ноги при возрастающем сопротивлении приводит к постепенному усилению потоотделения на ладонях.
В свете этих данных понятна обычно повышенная активность поперечнополосатой мускулатуры у невротиков в состоянии «покоя» и полезное влияние систематического обучения таких больных мышечному расслаблению [511]. С помощью психотерапии можно добиться снижения мышечного тонуса у таких больных, что создает благоприятные условия для поддержания гипота-ламической системы в условиях, более близких к нормальным. Якобсон учил своих больных расслаблять мышцы, не участвующие в тех или иных действиях («дифференцированное расслабление»). Глубокое воздействие «психических» факторов иллюстрируют эксперименты Эйффа [912, 913]. Простая арифметическая задача вызывала у нормальных лиц значительную активность, регистрируемую электромиографически, и повышение основного обмена на 36%, тогда как в тех же условиях, но при создании путем гипноза «созерцательного отношения» потенциалы действия практически отсутствовали, а интенсивность- обмена изменялась всего на 1%. Эта работа показывает, что состояние мышечного расслабления снижает реактивность гипоталамуса, а следовательно, и эмоциональную возбудимость. И наоборот, снижение при этом состоянии уровня центрального возбуждения (в результате психотерапии или гипноза) снижает нисходящие гипоталамо-соматические разряды, результатом чего должно быть постепенное снижение состояния возбуждения гипоталамуса вследствие снижения активности мышечных веретен в системе обратной связи.
Получены дополнительные данные о роли проприо-цептивиых импульсов в поддержании состояния возбуждения ретикулярной формации и, следовательно, косвенно — гипоталамуса и коры. При перерезке ствола под продолговатым мозгом (encephale isole [100]) в коре регистрируются асинхронные потенциалы, свидетельствующие о состоянии бодрствования. Перерезка соответствующих нервов, лишающая мозг притока зрительных,
,Ы| БИОЛОГИЧЕСКИЕ АСПЕКТЫ МЫШЕЧНОГО РАССЛАБЛЕНИЯ 395
слуховых*, вестибулярных и обонятельных раздражений и импульсации, передаваемой по блуждающему нерву, существенно не меняет этого состояния. Однако при двухстороннем удалении ядра V пары черепномозговых нервов в коре возникает синхронизация (как во время сна) [763]. Очевидно, импульсы от кожи и мышц, иннер-вируемых V парой нервов, играют особую роль в поддержании состояния бодрствования путем влияния на ретикулярную формацию. Учитывая тот факт, что для поддержания определенного уровня спинального возбуждения проприоцептивные импульсы имеют более важное значение, чем кожные [341], а также то, что активация гипоталамуса тесно связана с ретикулярной формацией, можно утверждать, что активность коры у только что описанных препаратов зависит главным образом, если не исключительно, от проприоцептивных импульсов, поступающих в мозг по V паре черепномозговых нервов.
ФАРМАКОЛОГИЧЕСКИЕ ИССЛЕДОВАНИЯ
Согласно данным фармакологических исследований, состояние мышечного расслабления может быть достигнуто введением панпарнита (мепанита). При измерении мышечной активности у иенаркотизированпого кролика было показано, что данное вещество снижает мышечный тонус как у нормального животного, так и у животного после перерезки спинного мозга. Порог полисинаптиче-ских рефлексов при этом не меняется. У человека наблюдалось снижение мышечного тонуса и возрастание порога проприоцептивного коленного рефлекса. Кроме того, ухудшалось восприятие весовых различий (измерявшихся по напряжению приложенной к руке пружины) [247а]. Эти данные подтверждают факт уменьшения притока проприоцептивных импульсов в результате расслабления мышц, вызванного введением препарата. То, что панпарнит способствует синхронизации в коре у препарата encephale isole и снижает пробуждающий эффект от раздражения гипоталамуса [595], вовсе не свидетельствует в пользу его центрального действия, поскольку, по имеющимся данным, влияние снижения проприо-
ГЛАВА XVII
цептивпой импульсации на состояние возбудимости гипоталамуса и гипоталамо-кортикальные разряды идентично депрессивному воздействию на ретикулярную формацию и гипоталамус.
С целью уменьшения эмоциональной реактивности у человека применялся мепробамат, причем было показано, что он снижает активность поперечнополосатой мускулатуры [191], хотя при этом не исключается влияние внушения [914]. В больших дозах он, видимо, устраняет повышение кровяного давления и ускорение сердечного ритма, возникшие под влиянием эмоционального возбуждения [914]. Тот факт, что мепробамат устраняет реакцию страха у человека [657] и снижает характерные (эмоциональные) реакции, наблюдаемые у крыс с разрушением промежуточного мозга [493], позволяет предположить, что это вещество снижает реактивность гипо-таламической системы, хотя еще не установлено относительное значение центрального депрессивного действия и снижения активности мышечных веретен. Следует указать, что во многих исследованиях применялись слишком высокие дозы, поэтому их нельзя использовать для интерпретации эффекта действия этого вещества на человека (см. [948]).
ДЕЙСТВИЕ ТЕПЛА И СОСТОЯНИЕ МЫШЕЧНОГО РАССЛАБЛЕНИЯ
Общеизвестно, что повышение температуры окружающей среды вызывает у человека и животных общее расслабление. Снижение мышечного напряжения и отсутствие движений вызывает уменьшение теплообразования в организме, что вместе с усилением теплоотдачи обеспечивает поддержание нормальной температуры тела. Установлено, что эта форма терморегуляции осуществляется благодаря повышенной активности переднего отдела гипоталамуса. При нагревании этой области наблюдается одышка, потоотделение и расширение сосудов, так что теплоотдача значительно возрастает [735]. Однако можно обнаружить и изменения более сложного порядка. При умеренном нагревании переднего отдела гипоталамуса снижается активность коры мозга.
ЕСКИЕ АСПЕКТЫ МЫШЕЧНОГО РАССЛАБЛЕНИЯ 397
Быстрые колебания ЭЭГ, характерные для состояния бодрствования, сменяются синхронизированными волнами и группами веретен, как при барбитуратном наркозе [915]. (Чрезмерное нагревание вызывает состояние возбуждения в коре, как при лихорадочном состоянии или локальном нагревании самой коры [888].)
В свете существования реципрокных отношений между передним и задним гипоталамусом можно предположить, что активность заднего отдела гипоталамуса, а следовательно, и тонические гипоталамо-кортикальные разряды снижаются и что этот сдвиг полностью или хотя бы отчасти лежит в основе упомянутых изменений корковых потенциалов. Кроме того, поскольку мы неоднократно подчеркивали, что задний отдел гипоталамуса играет важную роль в регуляции эрготропной системы, к которой относятся как симпатический отдел вегетативной нервной системы, так и поперечнополосатая мускулатура, регулируемая соматической нервной системой, можно ожидать, что реципрокное торможение заднего отдела гипоталамуса должно проявиться также в снижении тонуса скелетных мышц. Можно еще глубже проанализировать рассматриваемые явления, если исследовать изменение разрядов мышечного веретена (при постоянном напряжении мышцы) при повышении температуры переднего отдела гипоталамуса. Оказалось, что нагревание переднего отдела гипоталамуса сопровождается появлением медленных потенциалов в коре и в то же время — прекращением активности мышечных веретен [915]. Очевидно, нагревание переднего отдела гипоталамуса снижает центральное возбуждение промежуточного мозга, который, действуя через систему тонких гамма-волокон, определяет реактивность мышечных веретен и, следовательно, мышечный тонус. Поскольку пониженный мышечный тонус в свою очередь снижает возбудимость заднего отдела гипоталамуса, становится понятным, что с повышением температуры окружающей среды связано расслабление мышц, сонливость и ослабление эмоциональной реактивности. Медицина издавна использует благотворное действие теплых ванн.
Date: 2016-11-17; view: 239; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |