Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Ф- Конте, М. Грумбах
НАРУШЕНИЯ ПОЛОВОЙ ДИФФЕРЕНЦИРОВКИ
Благодаря успехам молекулярной генетики, экспериментальной эмбриологии, биохимии стероидных гормонов, а также новым методам изучения гипотала-мо-гипофизарно-гонадной системы стали во многом понятны механизмы определения пола и половой диф-ференцировки. Нарушения половой дифференцировки могут возникать на любой стадии внутриутробного развития и проявляться в разном возрасте — как сразу после рождения, так и по окончании полового развития.
НОРМАЛЬНАЯ ПОЛОВАЯ ДИФФЕРЕНЦИРОВКА
Генетический пол
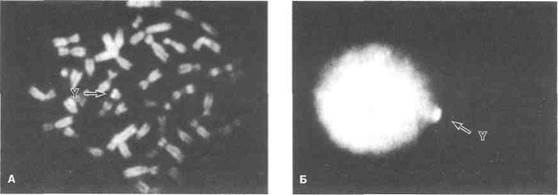
В норме соматические клетки человека содержат 22 пары аутосом и одну пару половых хромосом — XX или XY. Ка-риотип — это совокупность морфологических характеристик полного набора хромосом. Дифференциальное окрашивание позволяет по характерному набору полос идентифицировать каждую хромосому (рис. 44.1). При Q-окрашивании используют флюоресцентный краситель акрихин, при G-окрашивании — краситель Гимзы. С-окрашивание применяют для идентификации центро-мерных районов хромосом. Эти методы позволяют без труда выявить Y-хромосому по яркой флюоресценции как в метафазных, так и в интерфазных ядрах (рис. 44.2). Новый метод — флюоресцентная гибридизация in situ — особенно полезен для выявления так называемых маркерных хромосом, особенно половых хромосом с делениями, которые трудно обнаружить при стандартном окрашивании. В табл. 44.1 приведена номенклатура хромосом человека и их некоторых аномалий. Недавно был полностью картирован эухроматический район Y-хромосомы, образованный примерно 35 млн нуклеотидов.
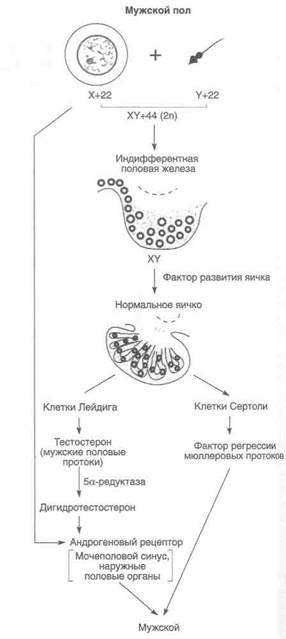
Как показали экспериментальные исследования и обследование больных с нарушениями половой дифференцировки, половые хромосомы и аутосомы несут гены, определяющие направление половой дифференцировки, то есть развитие из индифферентной половой железы яичка или яичника. Для развития яичников необходимы две нормальные Х-хромосомы и отсутствие Y-хромосомы (а также генов, определяющих развитие яичка). Для формирования яичек нужны Y-хромосома или район короткого плеча Y-хромосомы, кодирующий фактор развития яичка.
У человека Х- и Y-хромосомы значительно отличаются по размеру. Компенсация дозы генов при наличии двр и более Х-хромосом достигается частичной инактивацией всех Х-хромосом, кроме одной. Этот случай-
ный процесс наблюдается в каждой клетке на стадии поздней бластоцисты, когда происходит гетерохрома-тинизация материнской или отцовской Х-хромосомы. В результате образуется половой хроматин (тельца Барра), который обнаруживают в интерфазных ядрах, несущих две и более Х-хромосомы (рис. 44.3). Ген XIST находится в парацентромерном районе длинного плеча Х-хромосомы (Xql3.2), в предполагаемом центре инактивации Х-хромосомы. Предполагается, что с этого гена транскрибируется крупная РНК, которая каким-то образом связывается с Х-хромосомой и обеспечивает ее инактивацию.
Дистальный участок короткого плеча Х-хромосомы не инактивируется и включает короткий район (2,5 млн нуклеотидов), гомологичный району на дистальном участке короткого плеча Y-хромосомы. Этот район назван псев-доаутосомным. Именно псевдоаутосомные районы Х- и Y-хромосомы подвергаются конъюгации и кроссинго-веру во время мейоза, обеспечивая обмен генетическим материалом. В псевдоаутосомных районах Х- и Y-хромосомы обнаружено по крайней мере 7 генов. Среди них MIC2 (кодирует антиген CD99, распознаваемый мо-
Рисунок 44.1. Нормальный кариотип 46,XY. После окрашивания красителем Гимзы видны G-полосы. Каждой хромосоме присущ характерный набор полос. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
Глава 44. Нарушения половой дифференцировки
|
Рисунок 44.2. Флюоресцентная микроскопия (Q-окрашивание). Яркая флюоресценция позволяет обнаружить Y-хромосому как в метафазных, так и в интерфазных ядрах. А. Метафазные хромосомы. Б. Интерфазные хромосомы. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.

 ноклональным антителом 12Е7), ген гранулоцитар-но-макрофагального колониестимулирующего фактора, ген рецептора ИЛ-3 и ген низкорослости SHOX. Этот ген экспрессируется в костях. У гетерозигот его присутствие проявляется низкорослостью и дисхондростеозом. У гомозигот развивается мезомелическая дисплазия типа
ноклональным антителом 12Е7), ген гранулоцитар-но-макрофагального колониестимулирующего фактора, ген рецептора ИЛ-3 и ген низкорослости SHOX. Этот ген экспрессируется в костях. У гетерозигот его присутствие проявляется низкорослостью и дисхондростеозом. У гомозигот развивается мезомелическая дисплазия типа

Рисунок 44.3. Половой хроматин (тельце Барра) в ядре клетки слизистой щеки здоровой женщины (кариотип 46,ХХ). Gram-bach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
Лангера, для которой характерна диспропорциональная карликовость. Псевдоаутосомными районами являются также дистальные участки длинного плеча Х- и Y-хромо-сомы. В них обнаружен только один ген (рис. 44.4 и 44.5). В соскобах со слизистой щеки у здоровых женщин (кариотип 46,ХХ) в 20—30% интерфазных ядер обнаруживают половой хроматин, который отсутствует у здоровых мужчин (кариотип 46,XY). При полисомии по Х-хромосоме число телец Барра в диплоидном ядре на единицу меньше числа Х-хромосом. Подсчет телец Барра и Q-окрашивание хромосом используют для определения числа и типа половых хромосом (табл. 44.2).
Определение пола
За последние 20 лет изучено несколько белков, которые предполагались на роль фактора развития яичка, кодируемого геном на Y-хромосоме. Экспериментальные исследования и клинические наблюдения не подтвердили, что этим фактором являются антиген H-Y и белок, содержащий цинковые пальцы. При исследовании лиц с кариотипом 46,XX и мужским фенотипом и транслокацией небольшого участка с Y- на Х-хромосому был обнаружен новый ген, расположенный тотчас проксимально от псевдоаутосомного района на коротком мече Y-хромосомы (рис. 44.5). Этот ген был клонировани назван геном SR У (sex determining region Y — ген определения пола). У мышей ген Sry (аналог гена £ЛУчелове-ка) экспрессируется в половых тяжах между 10-ми и 12-ми сутками эмбриогенеза, то есть непосредственно перед развитием и в начале развития яичка. Делеции и точечные мутации гена SRY обнаруживают у 15% лице кариотипом 46,XY и женским фенотипом при чистой дисгенезии гонад. В том, что именно ген 57?Укодирует фактор развития яичка, больше всего убеждает то, что встраивание в геном клеток мышиных эмбрионов с кариотипом 40,ХХ гена Sry приводит к рождению самцов,
Глава 44. Нарушения половой дифференцировки
Таблица 44.1. Номенклатура хромосом человека и их некоторых аномалий
| Международная цитогенетическая номенклатура хромосом человека (ISCN, 1995) | Описание | Старая номенклатура | |
| 46,ХХ | Нормальный женский кариотип | XX | |
| 46.XY | Нормальный мужской кариотип | XY | |
| 47.XXY | Трисомия по половой хромосоме (47 хромосом, дополнительная Х-хромосома) | XXY | |
| 45,Х | Моносомия по Х-хромосоме | хо | |
| 45,X/46,XY | Мозаицизм (клетки с кариотипами 45,Х и 46,XY) | X0/XY | |
| Р | Короткое плечо хромосомы | Р | |
| Ч l,X,del(X)(p21) | Длинное плечо хромосомы Делеция короткого плеча Х-хромосомы дистально от полосы Хр21 | q ХХр- | |
| 46,X,del(X)(q21) | Делеция длинного плеча Х-хромосомы дистально от полосы Xq21 | XXq- | |
| |,X,i(XqlO) 46,X,r(X)(p22q25) 46,XY,der(7)t(Y;7)(ql1 | ;q!3) | Изохромосома по длинному плечу Х-хромосомы (qlO — центромерная полоса) Кольцевая Х-хромосома с разрывами по Хр22 и Хр25 Транслокация дистального флюоресцирующего участка Y-хромосомы на длинное плечо 7-й хромосомы | XXqi XXr 46,XYt(Yq-7q+) |
Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
Фактор развития яичка — ДНК-связывающий белок, содержащий домен из 80 аминокислот, схожий с доменами HMG-белков (high mobility gel proteins — белки, обладающие высокой подвижностью при электрофорезе в геле). Этот домен связывается с участком ДНК с последовательностью А/ТАСААТ, изгибая молекулу ДНК и влияя на транскрипцию других генов. Почти все известные на сегодняшний день мутации, обнаруженные улиц с кариотипом 46,XY и женским фенотипом при чистой дисгенезии гонад, затрагивают ген SRYna участке, кодирующем ДНК-связывающий домен фактора развития яичка.
Число генов, участвующих в развитии яичка, точно не известно (рис. 44.6, А). Гетерозиготные мутации и де-леции гена WT1 (ген нефробластомы 1,11р13) приводят к порокам развития мочевых путей и половых органов и нефробластоме. У мышей выключение гена Wtl приводит к апоптозу клеток метанефрогенной ткани и, следовательно, отсутствию почек и половых желез. Таким образом, являясь регулятором транскрипции, ген WT1 обеспечивает развитие мочевых путей и половых органов, действуя на самых ранних его этапах.
Ген FTZF1 кодирует внутриклеточный рецептор NR5A1 (так называемый стероидогенный фактор 1) и также участвует в регуляции транскрипции. Он экспрессируется в половых тяжах эмбрионов как мужского, так и женского пола, а также во всех тканях, где происходит синтез стероидных гормонов. Ген FTZF1 необходим для синтеза тестостерона, в клетках Сертоли регулирует экспрессию гена, кодирующего фактор регрессии мюллеровых протоков. Выключение гена Ftrfl у мышей приводит к апо-
птозу клеток половых тяжей. В результате у животных обоего пола отсутствуют половые железы и надпочечники. Таким образом, ген FTZF1 играет ключевую роль в формировании всех стероидпродуцирующих желез — надпочечников, яичек и яичников.
| Таблица 44.2.Определение числа и типа половых хро- | ||
| мосом по числу телец Барра и наличию Y-хромосом в | ||
| интерфазных | ядрах соматических клеток | |
| Половые хромосомы | Число в диплоидной соматической клетке | |
| Тельце Барра | Y-хромосома | |
| 45,Х | ||
| 46,ХХ | ||
| 46,XY | ||
| 47,ХХХ | ||
| 47,XXY | ||
| 47,XYY | ||
| 48,ХХХХ | ||
| 48,XXXY | ||
| 48,XXYY | ||
| 49,ХХХХХ | ||
| 49.XXXXY | ||
| 49,XXXYY |
а Число телец Барра в диплоидных соматических ядрах на единицу меньше числа Х-хромосом. Число флюоресцирующих Y-хромосом равно числу Y-хромосом в кариотипе. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
Глава 44. Нарушения половой дифференцировки

|
 Рисунок 44.4. Схема расположения G-полос и некоторых генов на Х-хромосоме. ГМ-КСФ — гранулоцитарно-макрофагальныи колониестимулирующий фактор. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
Рисунок 44.4. Схема расположения G-полос и некоторых генов на Х-хромосоме. ГМ-КСФ — гранулоцитарно-макрофагальныи колониестимулирующий фактор. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
 У лиц с дупликацией района короткого плеча Х-хромосомы Хр21 наблюдается чистая дисгенезия гонад 46,XY (женский фенотип при мужском кариотипе). Ген, расположенный в этом районе, был назван геном DAX1 (ген до-зозависимой смены пола). Этот ген кодирует внутриклеточный рецептор NR0B1. Делеции или мутации этого гена приводят к врожденной гипоплазии надпочечников с первичным гипогонадизмом. При этом половые железы у лиц с кариотипом 46,XY дифференцированы по мужскому типу. Между тем у мышей с кариотипом 40,XY дупликация гена Ahch (гомолог ВАХТ) при наличии слабых аллелей гена Sty приводит к дифференцировке половых желез по женскому типу. Дупликация данного гена у особей с кариотипом 40,XX не нарушает дифференцировку половых желез. Таким образом, именно дупликация гена
У лиц с дупликацией района короткого плеча Х-хромосомы Хр21 наблюдается чистая дисгенезия гонад 46,XY (женский фенотип при мужском кариотипе). Ген, расположенный в этом районе, был назван геном DAX1 (ген до-зозависимой смены пола). Этот ген кодирует внутриклеточный рецептор NR0B1. Делеции или мутации этого гена приводят к врожденной гипоплазии надпочечников с первичным гипогонадизмом. При этом половые железы у лиц с кариотипом 46,XY дифференцированы по мужскому типу. Между тем у мышей с кариотипом 40,XY дупликация гена Ahch (гомолог ВАХТ) при наличии слабых аллелей гена Sty приводит к дифференцировке половых желез по женскому типу. Дупликация данного гена у особей с кариотипом 40,XX не нарушает дифференцировку половых желез. Таким образом, именно дупликация гена
DAX1 обеспечивает дозозависимую смену пола у человека, Поскольку мутации гена Ahch, приводящие к потере его функции, не влияют на развитие яичников и фертиль-ность у самок мышей, считается, что кодируемый им внутриклеточный рецептор скорее играет роль фактора, подавляющего развитие яичек, нежели фактора, определяющего развитие яичников (рис. 44.6, Б).
Кампомелическая дисплазия — это заболевание, при котором наблюдаются пороки развития скелета и чистая дисгенезия гонад 46,XY. Оно развивается при мутациях гена SOX9 (17q24.3-q25.1). Этот ген кодирует один из HMG-белков, сходных с фактором развития яичка,» на 60% гомологичен гену SRY. Улиц с кариотипом 46.XY и делециями короткого плеча 9-й и длинного плеча 10-й хромосом наблюдается дисгенезия гонад и мух-
Глава 44. Нарушения половой дифференцировки

|
Рисунок 44.5. Схема расположения G-полос и некоторых генов на Y-хромосоме. ГМ-КСФ — гранулоцитарно-макрофагальный колониестимулирующий фактор. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.


 ской псевдогермафродитизм, что указывает на участие расположенных здесь генов в половой дифференциров-ке(рис. 44.6, Б).
ской псевдогермафродитизм, что указывает на участие расположенных здесь генов в половой дифференциров-ке(рис. 44.6, Б).
Дифференцировка половых желез
Половые железы остаются индифферентными до 42-х суток эмбриогенеза (длина эмбриона 12 мм). К этому времени из экстрагонадного источника — дорсальной энтодермы желточного мешка в них мигрируют 300—1300 первичных половых клеток. Эти крупные клетки — предшественники оогониев и сперматогониев — абсолютно необходимы для развития яичника, но не яичка. Под влиянием гена 57? Ги других генов, определяющих половую дифференцировку по мужскому типу (рис. 44.6, Б), на 43—50-е сутки эмбриогенеза формируется яичко. Клетки Лейдига появляются примерно к 60-м суткам, а мужские наружные половые органы становятся различимы на 65—77-е сутки эмбриогенеза.
Яичники развиваются гораздо позже — на 77—84-е сутки эмбриогенеза. В эти сроки, когда у мужских эмбрионов яички уже сформированы, большинство оогониев (образуются при митотическом делении первичных половых клеток) вступают в профазу I деления мейоза и превращаются в ооциты I порядка. Так начинается развитие яичника. На 90-е сутки эмбриогенеза становятся различимы примордиальные фолликулы — ооциты I порядка, окруженные одним слоем гранулезных клеток, лежащих на базальной мембране. В отличие от яичек яичники в эмбриогенезе не секретируют гормоны (рис. 44.7).
Дифференцировка половых протоков
Дифференцировка половых протоков схематично представлена на рис. 44.8. К 7-й неделе у эмбриона появля-
ются закладки мужских и женских половых протоков. Из мюллеровых протоков (если они не регрессируют) формируются маточные трубы, тело и шейка матки, а также большая часть влагалища. У мужского эмбриона вольфовы протоки дают начало придатку яичка, семя-выносящим и семявыбрасывающим протокам и семенным пузырькам. При наличии функционально активного яичка происходит регрессия мюллеровых протоков. Она обусловлена секрецией фактора регрессии мюллеровых протоков — димерного гликопротеида, который синтезируется клетками Сертоли. Этот фактор вызывает регрессию мюллерова протока только на той же стороне, где он синтезируется. Дифференцировку вольфо-вых протоков регулирует тестостерон, секретируемый яичками. При наличии яичника или функционально неактивного яичка вольфовы протоки дегенерируют, а мюллеровы протоки развиваются. Для синтеза тестостерона необходим ген FTZF1, продукт которого связывается с промоторами генов, кодирующих 20,22-десмо-лазу и 17а-гидроксилазу.
Ген фактора регрессии мюллеровых протоков находится на коротком плече 19-й хромосомы и кодирует белок, состоящий из 560 аминокислот. С-концевая последовательность этого белка гомологична последовательности трансформирующего фактора роста р и (3-субъединице ингибина и активина. Секреция фактора регрессии мюллеровых протоков продолжается в клетках Сертоли до 8—10 лет, поэтому он может служить маркером наличия клеток Сертоли. Экспрессию гена фактора регрессии мюллеровых протоков регулирует внутриклеточный рецептор NR5A1 — продукт гена FTZF1. Ген, кодирующий рецептор фактора регрессии мюллеровых протоков, был клонирован (12ql3). Этот рецептор принадлежит к семейству рецепторов типа II трансформирующего фактора роста р и сходных с ним белков.
Глава 44. Нарушения половой дифференцировки
Глава 44.
| ются по женскому типу (рис. 44.10). Для развития мужских наружных половых органов необходимы тестостерон и особенно его производное дигидротестостерон, образующийся под действием 5а-редуктазы. У мужских эмбрионов источником тестостерона являются клети Лейдига. Сначала они, вероятно, секретируют тестостерон автономно, затем под влиянием ХГ, еще позже -под влиянием ЛГ, секретируемого гипофизом плода. Дифференцировка наружных половых органов и мочеполового синуса по мужскому типу происходит поддей- |

Рисунок 44.6. Схема последовательного включения генов в развитии яичек и секреции гормонов при половой дифференцировв по мужскому типу. ФМРП — фактор регрессии мюллеровых протоков; DAX1 — ген, кодирующий внутриклеточный рецептор NR0B1, дупликация этого гена подавляет экспрессию SR Y и, следовательно, развитие яичек; FTZF1 — ген, кодирующий внутриклеточный рецептор NR5A1 (так называемый стероидогенный фактор 1); SOX9 — ген семейства Sox, кодирующий один из HMG-белков, сходных с фактором развития яичка; WT1 — ген нефробластомы 1. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
Дифференцировка наружных половых органов
Дифференцировка наружных половых органов схематично представлена на рис. 44.9. До 8-й недели эмбриогенеза наружные половые органы у эмбрионов обоего пола имеют одинаковое строение. Развитие женских наружных половых органов не зависит от наличия яичников, поэтому при наличии тяжевидных гонад или аге-незии гонад наружные половые органы дифференциру-
Пример!
гестацион!
возр
37—45 43—50
80 i
11—12 н
20—25 н
Рисунок 44.
цировку вс Textbook ofi
ствием ди
и тестосте
вым рецеп
плекс с вь:
фическим:
В итоге по;
которые ре
ку клеток.
новый рец
стке длинг
образом, д(
женных на
Как уже
наружных
стимуляци
Следует скг
вых органо
личии диги
геновых ре
рост полове
губно-мопи
го члена и jv
образовани
од в реализ.
дели эмбри
срастаются
стимуляци»
Причины н
ловых оргаи

|
Глава 44. Нарушения половой дифференцировки

|
Рисунок 44.7. Половая дифференцировка в эмбриогенезе. Обращает на себя внимание, что развитие яичка опережает дифферен-цировку всех остальных структур. Grumbach MM, Conte FA: Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology,9thed. Saunders, 1998.
ствием дигидротестостерона. Дигидротестостерон, как и тестостерон, связывается с внутриклеточным белковым рецептором клетки-мишени. Образовавшийся комплекс с высокой аффинностью связывается со специфическими участками ДНК, запуская транскрипцию. Витоге под влиянием андрогенов синтезируются белки, которые регулируют пролиферацию и дифференциров-ку клеток. Ген, кодирующий внутриклеточный андроге-новый рецептор, расположен в парацентромерном участке длинного плеча Х-хромосомы (рис. 44.4). Таким образом, действие андрогенов зависит о генов, расположенных на Х-хромосоме.
Как уже было сказано, для формирования мужских наружных половых органов необходима андрогенная стимуляция начиная с ранних стадий эмбриогенеза. Следует сказать, что дифференцировка наружных половых органов по мужскому типу возможна только при наличии дигидротестостерона и внутриклеточных андрогеновых рецепторов. Дигидротестостерон стимулирует рост полового бугорка, слияние мочеполовых складок и губно-мошоночных складок с формированием полового члена и мошонки. В то же время андрогены тормозят образование влагалища. Существует критический период в реализации эффектов андрогенов. После 12-й недели эмбриогенеза губно-мошоночные складки уже не срастаются, несмотря на выраженную андрогенную стимуляцию. Рост полового члена при этом происходит. Причины нарушения дифференцировки наружных подовых органов по мужскому типу: 1) нарушение синтеза
или секреции тестостерона или его превращения в дигидротестостерон, 2) дефицит или дефект андрогеновых рецепторов, 3) нарушение синтеза и действия фактора регрессии мюллеровых протоков. Вирилизация наружных половых органов у плодов женского пола возможна при избытке эндогенных или экзогенных андрогенов, особенно до 12-й недели эмбриогенеза.
Психосексуальное развитие
Психосексуальное развитие означает формирование психологического пола, который объединяет четыре понятия: 1) формирование половой идентичности, 2) поло-ролевое поведение, или модель социального поведения, отличающая женщину и мужчину в соответствии с установленными в данном обществе традициями, 3) половую ориентацию, то есть выбор полового партнера, 4) особенности мышления и восприятия.
В последние 30 лет господствовала теория, согласно которой половая идентичность формируется после рождения в результате взаимоотношений с окружающим миром. Недавно эта точка зрения была оспорена. Согласно работам ряда авторов, действие андрогенов во внутриутробном периоде и экспрессия генов, расположенных на Y-хромосоме, определяют половую идентичность у больных с наружными половыми органами промежуточного типа. Ранее предполагалось, что формирование половой идентичности происходит к возрасту 18—30 мес. Сейчас считается, что эта возрастная гра-



Рисунок 44.8. Дифференцировка половых протоков. А. Индифферентная стадия. Б. Развитие женских половых протоков. Остатки первичной почки и вольфовых протоков образуют придаток яичника, околояичник и гартнеров канал. В. Развитие мужских половых протоков до опускания яичек в мошонку. Единственное производное мюллерова протока — привесок яичка. Предстательная маточка формируется из мочеполового синуса. Conte FA, Grambach MM: Abnormalities of sexual
determination and differentiation. In: Greenspan FS, Strewler GJ (eds): Basic and Clinical Endocrinology, 5th ed. Appleton & Lange, 1 997.
Глава 44. Нарушения половой дифференцировки


|

|

Рисунок 44.9. Дифференцировка наружных половых органов. Grambach MM, Conte FA; Disorders of sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
 | |||
 |

|
|
| Глава 44. Нарушения половой дифференцировки |
| при нарушениях половой дифференцировки, необходимы дальнейшие исследования половой идентичности, полоролевого поведения и половой жизни больных с половыми органами промежуточного типа. До получения этих данных следует сказать, что, согласно имеющемуся опыту психосексуального воспитания большие наружными половыми органами промежуточного типа, основную роль в формировании половой идентичности |
Рисунок 44.10. Схематическое изображение этапов половой дифференцировки у человека. Grambach MM, Conte FA: Disordersof sex differentiation. In: Wilson JD et al. (eds): Williams Textbook of Endocrinology, 9th ed. Saunders, 1998.
ница сильно размыта. Это подтверждается следующим фактом. При наличии нарушений развития вторичных половых признаков, особенно при недостаточности 5а-редуктазы, смешанной дисгенезии гонад, недостаточности 17р-гидроксистероиддегидрогеназы, больные даже в период полового развития могут захотеть сменить пол с женского на мужской. Чтобы дать более гибкие рекомендации по психосексуальному воспитанию
Глава ¥
играют ] бенок, ] этого вь подкреп тительш половоп тельное] ных и их
НАРУ] ДИФ<3
Класс!
Нарушен зультате < определя мосомах носят ли] мужском ставлена ловые opi псевдогег яичников или муже] дифферет
Дисген каналы
Синдром
Синдром
чин перви
Для взрос.
небольши
спермия.1
болыпинст
щеки выяв
ты кариота
стика всех
фельтера -
Y-хромосо
случаи заб<
Как пок;
риотипа 47
ных новорс
тей препуб
яички,нещ
поведенчес
бальных на)
может остаг
отсталость,
ется редко. ]
признаки де
ное оволосе
развитие мь

|

|
Глава 44. Нарушения половой дифференцировки
играют выбор пола, в котором будет воспитываться ребенок, психологическое и социальное подкрепление этого выбора в грудном и детском возрасте, а также его подкрепление эндогенными или экзогенными (заместительная терапия) стероидными гормонами во время полового развития. Огромную роль играют также длительное наблюдение и психологическая поддержка больных и их родственников.
НАРУШЕНИЯ ПОЛОВОЙ ДИФФЕРЕНЦИРОВКИ
Классификация
Нарушения половой дифференцировки возникают в результате отклонений в ходе сложного процесса, который определяется генами, расположенными на половых хромосомах и аутосомах. К истинным гермафродитам относят лиц, у которых имеются и яичко, и яичник. При мужском псевдогермафродитизме половая железа представлена яичком, а половые протоки или наружные половые органы (или и те, и другие) женские. При женском псевдогермафродитизме половая железа представлена яичником, а наружные половые органы промежуточного или мужского типа. Классификация нарушений половой дифференцировки приведена в табл. 44.3.
Дисгенезия извитых семенных канальцев
Синдром Клайнфельтера
Синдром Клайнфельтера — одна из самых частых причин первичного гипогонадизма и бесплодия у мужчин. Для взрослых больных характерны мужской фенотип, небольшие плотные яички длиной менее 3 см и азооспермия. Часто наблюдается гинекомастия. Кариотип большинства больных 47,XXY: в соскобе со слизистой щеки выявляют тельце Барра. Описаны и другие варианты кариотипа, в том числе мозаицизм. Общая характеристика всех вариантов кариотипа при синдроме Клайнфельтера — по крайней мере две Х-хромосомы и одна Y-хромосома. Исключение составляют крайне редкие случаи заболевания с кариотипом 46,XX.
Как показали исследования, распространенность кариотипа 47,XXY составляет 1 на 1000 случайно отобранных новорожденных мальчиков. При обследовании детей препубертатного возраста отмечаются небольшие яички, непропорционально длинные ноги, психопатии и поведенческие расстройства. При исследовании вербальных навыков выявляют снижение IQ, хотя общий IQ может оставаться в пределах нормы. Тяжелая умственная отсталость, требующая специального обучения, встречается редко. После полового развития наблюдаются такие признаки дефицита андрогенов, как гинекомастия, скудное оволосение на лице и туловище, микропения, плохое развитие мышц и евнухоидное телосложение. Взрослые
мужчины с кариотипом 47,ХХУ обычно имеют высокий рост из-за непропорционально длинных ног (рост соответствует 75-му процентилю). Без лечения у взрослых больных повышен риск остеопороза. Кроме того, при синдроме Клайнфельтера повышен риск сахарного диабета, варикозного расширения вен ног, хронических заболеваний легких и рака молочной железы. Риск рака молочной железы в 20 раз выше, чем у здоровых мужчин. Часто наблюдается задержка полового развития. При синдроме Клайнфельтера повышен также риск экстрагонадных герминогенных опухолей, в том числе ЦНС и средостения. Эти опухоли могут секретировать ХГ и вызывать преждевременное половое развитие.
Патологические изменения в яичках прогрессируют и зависят от уровней гонадотропных гормонов. У взрослых больных наблюдаются гиалиноз и склероз извитых семенных канальцев, отсутствие или тяжелое угнетение сперматогенеза, а также узловая гиперплазия клеток Лейдига. Хотя гиалиноз извитых семенных канальцев обычно бывает диффузным, он выражен у разных больных и даже в разных участках яичка одного больного не одинаково. Характерна азооспермия, а фертильность возможна только при мозаицизме 46,XY/47,XXY.
В формировании кариотипа 47,XXY играет роль нарушение расхождения хромосом в I и во II делении мейо-за. В 53% случаев он обусловлен нарушением расхождения хромосом в I делении мейоза у отца, в 34% — в I делении мейоза у матери, в 9% — во II делении мейоза. Только у 3% больных мутация обусловлена нарушениями митоза зиготы.
Заподозрить синдром Клайнфельтера можно по характерному фенотипу и отклонениям уровней гормонов. Диагноз подтверждают при выявлении телец Барра в соскобе со слизистой щеки и кариотипа 47,XXY при исследовании крови, кожи или половых желез. По окончании полового развития в сыворотке и моче повышены уровни гонадотропных гормонов, особенно ФСГ. Уровни общего и свободного тестостерона в сыворотке снижены, экскреция тестостерона и эстрадиола замедлена, хотя уровень эстрадиола в сыворотке в норме или повышен. При биопсии яичка обнаруживают классическую гистологическую картину: гиалиноз извитых семенных канальцев, резкое уменьшение сперматогониев и узловую гиперплазию клеток Лейдига.
При синдроме Клайнфельтера проводят заместительную терапию андрогенами. Она особенно показана при задержке или остановке полового развития, а также при снижении уровня тестостерона в сыворотке. Тестостерон стимулирует развитие вторичных половых признаков и половую функцию, предотвращает остеопороз, предотвращает или уменьшает гинекомастию, а также улучшает самочувствие. При дефиците тестостерона подросткам назначают тестостерона энантат, 50 мг в/м каждые 4 нед, постепенно увеличивая дозу до 200 мг в/м каждые 2 нед. В более позднем возрасте для заместительной терапии можно использовать пластыри с тестостероном. Тестостерон бывает эффективным при умеренной
Глава 44. Нарушения половой дифференцировки
Глава 4Z
Таблица 44.3. Классификация нарушений половой дифференцировки
Date: 2016-11-17; view: 777; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |