Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
арапайым эволюциялық бірлік және қарапайым эволюциялық құбылыс туралы түсінік
Табиғатты кез келген жануар, өсімдік немесе ұсақ ағза түрлерінің даралары өз ареалының көлемдігі аймақты толық иемденбей, тек тіршілігіне қолайлы белгілі үлескілерді ғана мекендейді. Мысалы, бір түрге жататын бунақденелілердің ағаш жапырақтарымен қоректенетін даралары орман алқаптарында ғана, ал шөптекті өсімдіктермен қоректенетін даралары шалғындық үлескілерде мекендейді. Сол себепті түрдің даралары өз аймағының көлемінде түрлі тіршілік жағдайларына байланысты жиілігі әр түрлі топтар құрып бытырап кетеді. Олай болса, мұндай жекелеген табиғи топтар құрып бытырап кетеді. Олай болса мұндай жекелеген табиғи топтар популяциялары түзеді. Популяция дегеніміз: эволюциялық ұзақ уақыт бойы ареалдың белгілі бір бөлімінде тіршілік етіп, өз алдына дербес генетикалық жүйе құра алатын, еркін шағылысып, өсімтал ұрпақ беретін бір түр дараларының шағын тобы. Бұл анықтамадан популяцияны тіршілік әрекеттері эволюциялық заңдылықтарға сай жүретін даралардың ең кіші қарапайым тобы деп түсінуге болады. Ал табиғатта кездесетін шағын ұрпақтар тобында, мысалы, «тұқымтоп панмиксиялық бірлестіктер». Сондай-ақ кішігірім жай топтасуларда немесе жеке эволюция көлемінде олар өз уақыт аралығында ғана тіршілік етуіне байланысты эволюциялық өзгерістерге ұшырап үлгермей жайылып кетеді.
Популяция даралары оқшаулануға көршілес популяциямен салыстырғанда барлық қасиеттері мен белгілері бойынша өзара өте ұқсас болып кетеді. Сонымен қатар популяция ішіндегі жынысты әр түрлі даралардың бір-бірімен кездесу мүмкіндігі де едәуір жоғары болады.
Популяциялардың араласып кетуіне көптеген кедергілер тау, өзен, теңіз, құм т.б. биологиялық кедергілер – жануарлардың ұя, ін салу мерзімдері мен шағылысу кезіндегі мінездегі және аталық пен аналық құрлысындағы ерекшеліктері, ал өсімдіктерде гүлдеу, тозаңдану кезіндегі және аталық пен аналық құрылысындағы ерекшеліктер болып табылады. Популяция – эволюцияның негізгі өлшем бірлігі. Себебі барлық бастапқы эволюциялық әрекеттер популяция ішінде жүреді. Мәдени өсімдік іріктемелері, үй хайуанаттарының қолтұқымдары және ұсақ ағзалардың штаммдары – адамның қолымен жасалған популяциялар.
Популяциялық ареалдың көлеміне байланысты ондағы даралардың сан мөлшері де айтарлықтай өзгеріп отырады. Далалы жерлердегі бунақденелілер мен майда өсімдіктер популяцияларының сан мөлшері көбінесе жүз мыңдаған және миллиондаған даралардан құралады. Кез келген популяция табиғи сұрыпталудың нәтижесінде өзі тіршілік еткен ортаға бейімделетін болады. Ұзақ уақыт әсер еткен табиғи сұрыпталуға байланысты популяциядағы даралар біркелкі генотиптерге ие болуы тиіс. Бірақ шын мәнінде олай болмайды. Популяциядағы генотиптер жағынан гетерогенді болып келеді. С.С. Четвериковтың айтуынша суға қаныққан губка тәрізді популяция да алуан түрлі мутацияларға толы болады. Мұны ол дрозофиме шыбынының жабайы популяцияларына туыстық шағылыстыру жасай отырып дәлелдеді. Популяциядағы генетикалық гетрогендікті С.С. Четвериков былай түсіндіреді: кез келген түр немесе популяциядағы даралар тіршілік барысында бірнеше ұрпақ бойы үздіксіз мутантты гендермен толықтырып отырады. Сонымен қатар басқа популяциядан ауысып келетін даралардың шағылысуы арқылы жаңа генетикалық материалдың популяцияға енуі байқалады. Мутациялардың көпшілігі әдетте рецессивті болады. Сондықтан сан мөлшері жоғары популяцияларда мұндай рецессивті мутациялар гетрозиготалық жағдайда тіпті байқалмайды. Құрамындағы даралардың гетрогендігіне қарамастан, кез келген популяция динамикалық тепе-теңдікте тұрған күрделі генетикалық жүйе болып табылады. Популяциядағы гендер артқан сайын гетрозиготалы ағзалардың өзара шағылысуы мүмкіндігі де артады. Соның нәтижесінде даралардың бірқатары гомозиготалы жағдайға көшіп, ұрпақтар арасында мутациялық өзгерістер айқын көрініс береді. Олардың ішінде ағаның тіршілік қабілетін төмендететін мутациялардың да кездесуі ықтимал. Бірақ табиғи популяцияларда генетикалық гетрогендік жеткілікті дәрежеде болғандықтан, популяцияның қалыпты тіршілігі ұзақ уақыт бойы сақталады. Қорыта келгенде популяция – кішігірім, дербес эволюциялық құрылым. Сондықтан табиғатта ақиқат тіршілік етіп, өзіне тән экологиялық морфофизиологиялық және генетикалық қасиеттерге ие болған популяция – эволюцияның қарапайым өлшем бірлігі болып табылады.
11. Харди-Вайнберг теңдеуі және оның популяция гендері мен генотиптерінің жиілігін анықтау үшін қолданылуы.
Популяциялық генетиканың негізін Харди-Вайнберг заңы құрайды. Тұқым қауалаушылықтың корпускулдық теориясы жеке генетикалық факторлар ата-аналардан ұрпақгарға өзгеріссіз беріледі деп түсіндіреді. Осыны 1908 ж. ағылшьн математигі Дж-Харди және неміс генетигі Г.Вайнберг бір-біріне тәуелсіз дәлелдеді.
Харди-Вайнберг заңы мынадай жағдайларды керек етеді:
— популяцияға кіретін организмдер саны жеткілікті мөлшіерде көп болуын;
— әр түрлі генотиптердің көбею қабіеттілігі бірдей болуын;
— популяцияда кездейсоқ шағылысудың сақталуын;
Харди-Вайнберг заңы формула арқылы өрнектеледі:
Айталық белгілі бір популяциядағы екі - А жөне а - аллельдің жиілігі р және q-re тең делік (p+q=I). А аллелі бар гаметалар р жиілікпен кездеседі, ал а аллелі бар гаметалардың жиілігі — q. Гаметалардың зиготаға қосылуы кездейсоқ өтетін болса, онда генотипі АА зиготаның жиілігі р2- қа, генотипі Аа зиготаның жиілігі 2pq- гежөне генотипі аа зиготаның жиілігі q2 -қа яғни биномдық формуланың жіктелу мүшелеріне (pA+qa)2 =p2AA+2pqAa+q2aa) тең болады.
Популяцияның бастапқы аталық және аналық дарақтарында аллельдер жиілігі бірдей болса, онда кездейсоқ шағылысуда кез келген генотиптерінің тепе- теңдік жиілігі бір ұрпақ көлеміңде іске асады.
12. Оқшаулану - қарапайым эволюциялық фактор ретінде. Оқшаулану құбылыстарының жіктелуі, оның эволюциялық рөлі.
Оқшаулану - панмиксияны бұзатын кез келген тосқауылдың пайда болуы. Эволюция процесіндегі оқшауланудың маңызы-еркін шағылыса алмау, ол популяция мен түрдің мекендеуші жеке бөліктерінің арасында айырмашылықтың бекуіне әкелді. Эволюциялық айырмашылықтың осындай бекітілуінсіз ешқандай форма түзіліс мүмкін емес. Оқшаулану кеңістіктік және биологиялық деп жіктеледі. Кеңістіктік оқшаулану әртүрлі формада болады: су тосқауылдары құрғақты мекендейтін түрлерді бөлсе, құрлық тосқауылдары гидробионт түрлерді оқшаулайды ал тауарлар жазықтың популяцияларды оқшаулайды. Осындай оқшаулану мысалына-салыстырмалы түрде аз жылжитын жануарлар-жербетілік моллюскалар-Гавай аралдарындағы кішкентай жазықшамен тіршілік етуге жарамды басқа жазықшамен оқшауланған; әрбір жазықтықта спецификалық ерекшеліктері бар өзіндік жеке популяциялар пайда болады. Осы сияқты территориялды-механикалық оқшауланудың пайда болуы түрлердің белгілі бір территорияда тариихи дамуына байланысты. Осы жағдайды оқшауланудың ең басты себебі мұз басу болып табылады. Қазіргі уақытта биосфераға адам әсер етуімен осындай кеңістіктік оқшаулану жиі болуда. Мысалға соболь (Martes zibellina) ХХ ғасырдың басында Еуропада ажыраған ареалы пайда болған. Әдетте, осындай ажыраған ареалдың пайда болуы-түрдің біртіндеп жоғалуының қауіпті белгісі болып табылады.
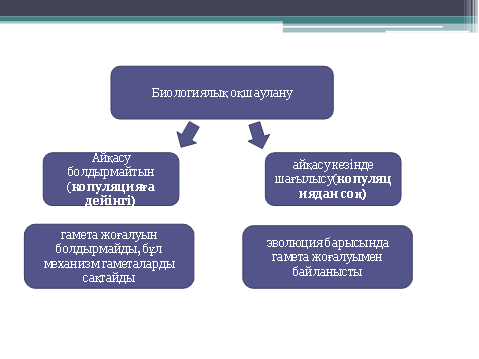
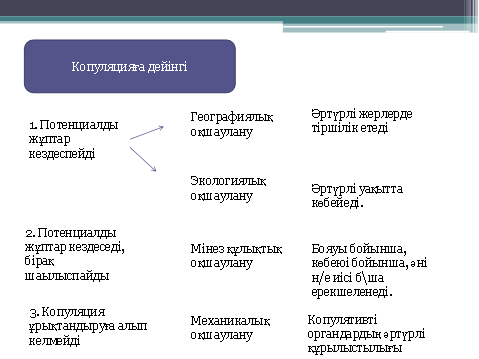

Географиялық оқшаулану. Жұмақтай сауысқандар Жаңа Гвинеяның тропикалық ормандарында мекендейді. Бес түрдің біреуі өзінің таулы аймағында қоректенеді. Түрлер арасындағы морфологиялық айырмашылық өте аз, кейде оларды бір түрге жатықызып та жатады: Черногорлая астрапия, Великолепная астрапия, Астрапия принцессы Стефании. Экологиялық оқшаулану. Тан көліндегі барбус балықтарының жақын туыстары шоғырланған. Көлде басқа балық түрлері аз болғандықтан, барбус балықтары барлық экологиялық ареалды игеріп алған.
13. Түр генетикалық тұрақты жүйе екендігі туралы түсінік. түр түзілудің негізгі жолдары. Аллопатриялық және симпатриялық түртүзілу. Олардың тәсілдері. Аллопатриялық түр түзілу. Түр түзілудің ең негізгі екі түрі бар: Аллопатриялық және симпатриялық түр түзілу.Аллопатриялық түрлердің түзілу процесі әдетте олардың бастапқы түрінің ареалының өзгеруіне байланысты. Бұл өзгерістер екі сипатта болуы мүмкін: түр өзінің ареалын кеңітіп, жаңа территорияны иемденеді, немесе физикалық кедергілердің ықпалымен түрдің ареалы бөлшектеніп кетіп, жеке популяциялар бөлініп оқшау қалып қояды. Бұл екі жағдайда да жаңа түр тармақтары пайда болуы арқылы келіп жаңа түр түзіледі.Түр ареалының қаншалықты өзгеруі мүмкін екендігін көрсету үшін мынандай мысалдар келтіруге болады. 1895 жыла дейін Москва және Тула облыстарының территориясынан жасыл түбіртек торғай өте сирек кездесетін. 1910 жылы ол Смоленск облысының территориясында кездесті, ал қазір Одерда ұялады.Ареалдың бөлшектенуі қашан да болса әр түрлі тарихи себептерге байланысты түрліше болуы мүмкін. Мысалы, тұтас ареалдың бірқатар бөлігінде климат жағдайларының өзгеруі және ондағы осы түрдің популяциялары жойылып кету салдарынанареал бөлшектенеді. Климат жағдайларының өзгеруіне байланысты түр өзінің ескі ареалының көлемінде көшіп қоныс аударуы мүмкін. Мысалы, мұздық дәуірде Европадағы альпілік ескі фауна мен флора таудан төмен жылжып түсіп, басын мұз басқан Альпі мен Солтүстіктен жылжып келе жатқан мұздықтың аралығындағы қыспақта орналасты. Мұздық дәуір өткеннен кейін өсімдіктер мен жануарлардың суыққа төзімді түрлері бұрынғы бағытына керісінше қоныс аударды: организмдердің бірқатары таудың басына қарай, бір қатары солтүстікке қарай шегініп бара жатқан мұздықтың ізімен жылжып отырды. Орталық Европа жазықтықтарындағы тіршілік жағдайлары альпілік организмдерге қолайсыз болғандықтан, альпілік түрлердің ареалдары бөлшектеніп кетті. Құрлықтың лықсып төмен түсуі немесе көтерілуінен, су қоймалары құрылысының өзгеруінен басқада себептерден ареалдар бөлшектеніп кетуі мүмкін.Түрдің ареалы неғұрлым көлемді болса, соғұрлым ондағы орта жағдайлары әр қилы келеді, яғни ареалдың экологиялық бөлшектенуі неғұрлым айқын болса, соғұрлым жеке популяциялардың не географиялық расалардың (түр тармақтарының) оқшауланып қалу мүмкіндігі мол болады. Экологиялық бөлшектенген ортада түр тармақтарын қалыптастыратын микроэволюциялық процесс күшті жүреді.Географиялық расалардың көбі түр мен түр тармақтарының шекарасында тұр. Бұл оқшауланған түр тармағынан жаңа түр түзілу процесін түсіну үшін өте қажетті факт болып саналады. Ауыспалы түр тармақтарының өз ара бір-бірімен жалғасып жатқан тізбегі шеңбер түзеп, оның екі шеткі ұштары түйісіп тұйықталып отырады. Бірақ, осы шеңбердің екі жағындағы мүшелері бір-бірінен алшақтап кеткендігі соншалықты, бұлар бір-бірімен буындаспайды, яғни бұлар өз алдына оқшау түр бейнесінде болады. Симпатриялық түр түзілу. Бір туысқа қарасты түрлердің ареалдары мүлдем оқшау болуы да, немесе олар бір-біріне жанасып, іргелес жатуы да, тіпті тұтас та болуы мүмкін. Оқшау ареалдары бар түрлер аллопатрикалық түрлер делінеді, іргелес немесе тұтас ареалдағы түрлер симпатрикалық түрлер делінеді. Бұл түрлердің ареалдары өзінше ерекше: бұлар үздік-үздік шеңбер бейнесінде Австралияның орталық бөлімін айнала қоршап жатып. Таяуда Австралияның климаты онша құрғақ болмағандығын және орман оның орталығына дейін еніп жатқандығын сене алсаң аталған ареалдардың арғы тегі түсінікті болады. Құрғақшылық кезеңнің шабулы орман алқаптарын ығыстырып азайтты, Platycercus туысындағы түрлердің қалыптасуы осымен байланысты. Симпатрикалық түрлермен істес болған жағдайда бұлардың түрлік ерекшелігі экологиялық ерекшеліктерін зерттеу нәтижесінде анықталады. Бұл ойды мынандай мысалмен түсіндіруге болады. Совет Одағының Европалық бөлімінің орта өңірінде сарғалдақтардың бір-бірінен таяу өсіп тұрған бірнеше түрлерін кездестіреміз: кашуби, көп гүлді (R. polyanthemus), күйдіргіш (R. acris), жатаған (R. repens) және безеулі (R. flammula) сарғалдақтар. Марфологиялық критерийді қолданғанда, бұларда айырмашылық бар екені көрінеді. Мысалы, кашуби сарғалдағының сабағының төменгі жағында ірі, бүйрек бейнелес жапырағы болады, осыған қарап оны ажырату оңай. Жапырағына қарай безеулі сарғалдақ та тез ажыратылады: оның жапырақтары ұсақ, сопақ, майда тісті жиекті болып келеді. Қалған үш түрден тек жатаған сарғалдақта ғана төселмелі ұзын өркендер болады. Көп гүлді сарғалдақ бір-біріне өте ұқсас. Бұлардағы айырмашылық мұның біріншісінде сабағының төменгі жағындағы жапырақтары жіңішке бөлімдерге терең тілімденген, гүл таянышында жүлгелері бар, екіншісінің жапырақтары бес бұрышты, саусақ-тілімді, ұзын сағақты; гүл таянышында жүлгелері болмайды.Осы түрлердің мекендеген биотоптарының ерекшеліктерін зерттеу: бұлар географиялық бір орында өсе тұрса да, әрқайсысы табиғат жағдайларының белгілі бір жиынтығын мекен ететіндігін көрсетеді. Сөйтіп, осы түрлердің әрқайсысына экологиялық ерекшеліктердің сыры ашылып отыр. Аталып өткен критерийлер бір-бірін толықтыра отырып, түрге тән ерекшелікті ашуға көмектеседі. Сондықтан да, жануарлар мен өсімдіктердің түрлерін сипаттағанда, олардың марфологиясының, физиологиясының және экологиясының қай жақтары қандай дәрежеде зерттелуіне байланысты осы критерийлердің әрқайсысы түрге тән қандайда бір ерекшеліктерді анықтауға көмектеседі. Марфологиялық талдаудың өзі-ақ біздің алдымыздағы үш түрлі органикалық формалар екендігін көрсетеді. Генетикалық және физиологиялық ерекшеліктері де бұладың дербес форма екенін дәлелдейді. Сөйтіп, біздің алдымызда марфология, генетика, физиология және экология жөнінен үш түр – оқшау органикалық форма, яғни үш дербес түр тұрғанынакүмәндануға болмайды. Бұл түрлер түрліше ортада қалыптасқан болуға тиіс: біреуі шөлейт зонасында, екіншісі – далалықта, ал үшіншісі ормандық зонада.Аталған критерийлер организмдерді барлық топтарына қолданыла тұрса да, топ көлеміндегі биологиялық формалардың құрылыстарының және тіршілік бағытттарының ерекшеліктеріне байланысты әр түрлі таксондардың әрқайсысының өзіне тән ерекшелігі бар. Кейбір өсімдіктер мен жануарлардың топтарындағы түрлерді анықтау үшін олардың жеке органдарын истологиялық талдау жасау арқылы ғана марфологиялық критерий қолдануға болады.Жануарлардағы этиологиялық фанторлармен тығыз байланысты оқшауланушы механизмдерді зерттеуде түр критерийін қолданудың өзінше ерекшелігі бар. әр түрлі құрылыс дәрежесіндегі жануарлардың мінез-құлықтарының қалпы түрліше болуына байланысты, өсімдіктерге қарағанда бұлардың түр ішіндегі өзара қарым-қатынастары өте күрделі және алуан түрлі болады.
Түр - жүйе ретінде. Н.И Вавилов осы ұғымды қолдана отырып өсімдікке жұмыс жасағанымен, бұл концепция – жануарларда да қолданады. Бұл концепцияның басты идеясы – түр түзілістің проблемасына жақындау болып табылады. Вавилвтың айтуынша түрді құрылымға бөлу керек. Мысалы: түр→ түрасты т.б. Вавилов – Жорданонды мойындамады. Ал Джордонон – түрлер 1-ақ белгісі бойынша ажыратылатынын айтқан болатын. Түр – еркін шағылыса алатын және өсімталды ұрпақ беретін, белгілі орта жағдайларына бейімделген және арнайы ареалы бар, ұзақ уақыт бойы тіршілік ете алатын, морфофизиологиялық жағынан ұқсас дарабастар жиынтығы.
14. Филогенездің негізгі формалары: филетикалық эволюция, дивергенция, конвергенция, параллелизм. Топтардың дамуының қарапайым түрі. Даму барысында бұрын біртұтас болған нәрсенің екі немесе бірнеше түрге бөлінуі (дивергенция) және оның біртіндеп өзгеріп, өзге түрге айналуы жүреді (филетикалық даму). Филетикалық даму – даму барысында ағаштың кез келген діңнің немесе бұтағын бастапқы түрінен өзгертудің жүзеге асыратын міндетті процесс. Түрдің өзгеруі өзгеше түрді құрайтын кескінге тиіп өтуі мүмкін, оған тимеуі де мүмкін немесе аз ғана деңгейде (егіздер түрі), тек ішкі өзгеоістеріне тиюі мүмкін. ерттелген өмір ағымының палеоитологиялық діңдерінің көпмүшелігі филетиналық даму мысалын береді. Филетикалық дамудың типтік мысалы, жылқы тұқымдасының мына желі бойынша дамуы – Жирокотерум Эпигиппус – Мизогинпус – Парагиппус – Меригипус. Бір таксиологиялық топқа қатысты бір филогенетикалық діңдегі өзгеріс филетикалық даму мәнін құрайды. Мұндай өзгеріссіз ешқандай эволюциялық процесс өтпейді, сондықтан филетикалық дамуды дамудың қарапайым түрлерінің бірі деуге болады. Қандай да топтың филетикалық дамуы макроэволюциялық деңгейде оларды құрайтын түрлердің өзгерулерімен құралады (әдетте, ол туыстық торына кірмейді). Көзге әрбір талшығы үзілмей және қасынлдағысына қосылмай басынан аяғынадейін созылып тұрған көп қабатты арқан түрінде көрінеді. Макродеңгейдегі филетикалық өзгерулер қайтарымсыз; макродеңгейде будандасу кезінде жойылуы мүмкін. Дивергенция – топ дамуының басқа негізгі қарапайым формасы. Түрлі жағдайларда таңдау бағытының өзгеру нәтижесінде өмір ағашы бұтағының дивергенциясы (бөлінуі) жүреді. Органикалық көптүрліліктің және «өмір сомасының» тұрақты өсуі пайда болуының негізгі жолы осындай. Дивергенцияның бастапқы кезеңін түр-тұқымдық орналасуының жекелеген бөлігінде қандайда бір белгілер бойынша терең айырмашылықтар туындаған кезде түрдің ішкі деңгейін де байқауға болады. Популяция алшақтығы (бөлінуі) дамушы формалардың репродуктивті оқшаулауына, содан кейін түрленуіне алып келеді. Эволюцияның дивергентті формасы – макроэволюциялық топтардың түп негізі. Дивергентті эволюция механизмі түр ішіндегі эволюциялық факторларға байланысты. Изоляция нәтижесінде, өмір толқындары, негізінен мутивациялық процесте жинақтағанда популяция топтары эволюцияда белгілерін сақтап қалады, түрдің негізгі бөлігінен айқын ажыратылады. Эволюциядағы бір мезетте жиналған әр түрлілік бастапқы түрлердің жойылуына әкелуі мүмкін. Осыған ұқсас процестер дивергенция түрінде де байқалады. Эволюция барысында түрдің біреуінде маңызды иілімдер пайда болып басқа түрлерге шашырауы мүмкін. Әр түрлі топтар туыс аймағында дивергенция процесінде жаңа туыстың басталуына жол беруі мүмкін. Дивергенция кез келген масштабтың негізгі жинақтау іс-әрекетінің нәтижесі. Бірақ топтың жинақтау әр бірден кейін популяция ішіндегі жинақтауға негізделген. Түр ішілік дивергенция процестерінің ұқсастығына қарамастан, күрделі топтарға да, түрге қарағанда, бұл процестердің арасында айқын айырмашылық бар, микроэволюциялық деңгейге дивергенция процессінің қайтарылуы негізделген: екі ажырап қалған популяция қиылысу жолымен оңай қосыла алады және бір популяция түрінде қала береді. Дивергенция процестері макроэволюциялық қайтарылмайды. Дивергенция және диеллетикалық эволюция – филегенетикалық бұтақтың барлық өзгертулерінің негізі және табиғаттағы кез келген масштабтың эволюция процесінде элементарлы форманың ағысын айтады. Конвергенция және параллелизм. Дивергенциямен филетикалық эволюциямен салыстырғанда конвергенция және параллелизм эволюция топтарының ішінде өте күрделі болып табылады. Параллелизм (паралельді даму) – ұқсас екі немесе бірнеше біріншілік дивергенциялық топтардың филетикалық дамуындағы процесс. Белгілердің конвергенциясы. Егер екі түрлі, дегенмен жақын туыстарға жататын екі түрдің әрқайсысы өз кезегінде көптеген жаңа және өзара ажыратылатын формалар өндірсе, онда осы формалардың кейбіреулерінің бір-бірімен тығыз жақындасулары сонша, оларды жалпы бір туысқа кіргізуге тура келер еді, сондықтанда әр түрлі екі туыстың ұрпақтары біреу болып бірігер еді. Бірақ, егер біз бірінен бірі өте қашық тұрған формалардың өзгерген ұрпақтарындағы құрылыстың жалпы және жақын ұқсастығын конверген-цияға жатқызуды ойласақ, бұл өте ағаттық болар еді. Тірі организмдер жөнінде біз, олардың әрқайсысының формасы шексіз күрделі қатынастардан тәуелді екенін есте сақтауымыз қажет. Алайда бастапқыда өзара анық өзгешеленетін екі организмдердің ұрпақтарының барлық ұйымы толық ұқсас дәрежеге дейін жақындай алуы мүмкін емес. Егер бұл осылай болса, онда біз оның генетикалық байланыстарын тәуелсіз, сол форманы бірінен бірі қашық тұрған, қайталанушы геологиялық формациялардан кездестірер едік; бірақ геологияның барлық мәліметтері осыңдай жорамалға қарсы күәландырады. Тек бір ғана бей органикалық жағдайлар жөніне келсек, онда түрлердің жеткілікті саны, өзара өзгешеленетін жылылық, ылғалдылық және с.с. бар-лық жағдайларға едәуір жылдам бейімделген болып шығуы мүмкін секілді, бірақ мен организмдердің арасындағы өзара қатынастар өте маңызды рөл атқарады деп ойлаймын; кез келген елде уақыт өткен сайын түрлердің саны еседі, онда тіршіліктің органикалық жағдайлары да барған сайын күрделі бола береді. Осыдан бір қарағанда, құрылысындағы пайдалы әр түрліліктің кәлемінде шек жоқ, демек, пайда бола алатын түрлердің санында да шек жоқ болып көрінетін сияқты. Түрлердің санының шексіз өсуіне не кедергі жасайды? Тіршіліктің жалпы жиынын (мен мұнда түрлік формалардың санын ойламаймын) ұстап тұра алатын белгілі алаңның шегі болуы қажет, өйткені ол едәуір дәрежеде физикалық жағдайлардан тәуелді; осыдан, егер бұл алаң түрлердің аса көп санымен қоныстанған болса, онда әрқайсысы, немесе олардың әрқайсысы дерлік особьтардың тек азырақтау санынан тұратын болады, ал мұндай түрлер климат жағдайларының кездейсоқ құбылуының немесе олардың жауларының санының артуы нәтижесінде жойылуға ұшырайтын болады. Мұндай жағдайларда құру процесі тез жүруі қажет, сол кезде жаңа түрлердің түзілуі әрқашан өте баяу болады. Мынандай шекті жағдайды еске алыңыз, айталық, Англияда қанша особь болса, сонша түрлер болды делік, онда бірінші қатты қыс немесе құрғақ жаз көп мыңдаған түрлерді қырған болар еді. Сирек түрлер, – ал олардың саны шексіз өсу жағдайында барлық түрлер сирек болып шығады, – әлденеше рет түсіндірілген принцип бойынша, белгілі кезеңдердің шеңберінде пайдалы өзгерістер аздап түзеді; осыдан жаңа түрлердің пайда болу процесінің өзі тоқталып қалады. Қандай да болмасын түр өте сирек бола бастағанда, жақын дәрежелі туыстардың шағылысуы оны құртуға көмектеседі; кейбір авторлардың пікірі бойынша Литвадағы Зубрдың, Шотландиядағы қызыл бұғының, Норвегиядағы аюдың және басқаларының азаюын түсіндіретін себеп осы деп есептелінеді. Ақыр аяғында, – өз облысында көптеген бәсекелестерін жеңген басым түр, әрі қарай әлі де таралуға және басқа формаларды ығыстыруға тырысады – міне бұл ең басты бағыт деп ойлаймын. Д-р Гукер таяуда, Австралияның оңтүстік-шығыс бұрышында, жердің әр түрлі түкпірлерінен келушілердің көптігінен, жергілікті австралиялық түрлердің санының едәуір азайып кеткендігін көрсетті. Осы пікірлердің мәні қандай екенін айтқым келмейді, бірақ әр елде осы жағдайлардың бәрі жинақтала отырып, түрлік формалар санының шексіз өсу тенденциясын шектеуі қажет.
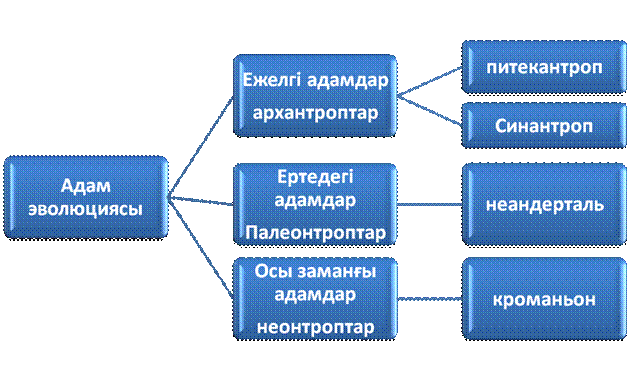
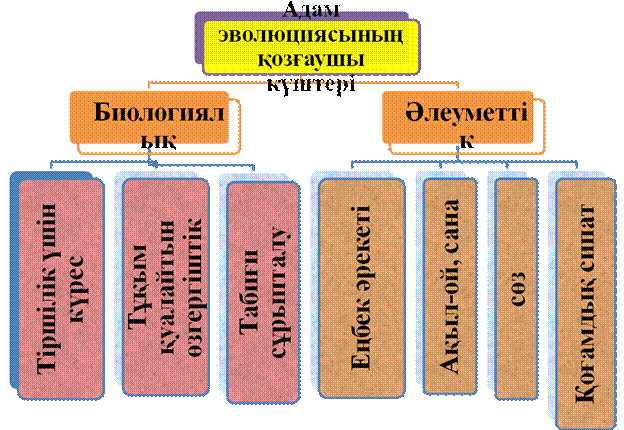
15. Адамның жануарлар дүниесін жүйелеудегі орны.
Антропогенез - (грек. anthropos – адам, genesіs – шығу тегі) – антропология ғылымының адамның шығу тегін, даму тарихын, оның жеке биол. түр болып қалыптасуын және адамзат қоғамының даму кезеңдерін әрі жаратылыстану, әрі қоғамдық ғылымдарға сүйене отырып зерттейтін негізгі саласы. Бұларды шешуде антропогенез приматология, эмбриология, физиология, психология, геология, археология, этнография, тіл білімі сияқты ғылымдардың нақты деректеріне сүйенеді.

Date: 2015-09-22; view: 4332; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |