Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Биологические объекты являются открытыми термодинамическими системами. Они обмениваются с окружающей средой энергией и веществом
Поддерживая внутренний порядок за счет энергии питательных веществ, любой организм всегда «выбрасывает» неупорядоченную энергию в окружающую среду в виде тепла или неусвоенных остатков пищи. Энтропия организма уменьшается, а
Энтропия окружающей среды увеличивается. При этом оказывается, что полная энтропия системы животное + окружающая среда увеличивается (внешний беспорядок превышает внутренний порядок). ∆S=∆So+∆Sc>0
В сложных биохимических реакциях, протекающих в клетке, законы термодинамики не выполняются. Пример. С молекулы ДНК постоянно списывается часть информации на и-РНК (транскрипция) для синтеза того или иного белка (трансляция). Этот белок синтезируется на рибосомах методом ускоренной конвейерной сборки, так, что скорость соединения аминокислот в белковую цепочку в десять раз превышает скорость пулементной ленты. Любые ошибки синтеза белка почти мгновенно исправляются контролирующими ферментами – тоже белками, которые синтезируются в той же клетке и тоже под контролем.
На этом «конвейере» нет ни одного лишнего движения. Всё это происходит при комнатных температурах, когда молекулы жидкостей и твердых тел колеблются с амплитудами, превышающими их размеры. При таких колебаниях биосинтез невозможен (всё равно, что попасть в игольное ушко трясущимися руками). Ученые установили, что в момент образования пептидной связи тепловые движения атомов прекращаются («замораживаются»), что и обеспечивает стопроцентную точность сборки. Это относится и ко всем внутриклеточным реакциям подобной сложности.
Строжайший и точнейший порядок частиц в организме рождается не от беспорядка, а от прежде установленного порядка, то есть все молекулярные процессы живого организма изначально запрограммированы.
- Основные типы транспорта вещества через биологические мембраны
Динамические свойства белков.
1. Латеральная диффузия. все значения для белков с М=100000, К=3*10–10 см/сек. Но белки могут объединяться в кластеры, которые мало подвижны.
2. Вращательная диффузия К=0,34 сек.
3. flip-flop переходы, К=10–4 сек – частота flip-flop перехода.
Большинство процессов жизнедеятельности, таких, как всасывание, выделение, проведение нервного импульса, мышечное сокращение, синтез АТФ, поддержание постоянства ионного состава и содержания воды связано с переносом веществ через мембраны. Этот процесс в биологических системах получил название транспорта.
Если перенос вещества происходит с уменьшением электрохимического потенциала, то есть не требует затрат энергии, то такой транспорт называется пассивным. Его разновидностями являются диффузия (перемещение веществ в сторону меньшей концентрации) и фильтрация (просачивание веществ поры в сторону меньших значений давления). С помощью диффузии в клетку проникают растворенные молекулы кислорода и углекислого газа, а также яды и лекарственные препараты. Примером фильтрации в организме является перенос воды через стенки кровеносных сосудов, выдавливание плазмы крови в почечные канальцы.
Транспорт веществ через липидный бислой с помощью простой диффузии совершается с малой скоростью, особенно в случае заряженных частиц, и почти не контролируется. Поэтому в процессе эволюции для некоторых веществ появились специфические мембранные каналы и мембранные переносчики, которые способствуют повышению скорости переноса и, кроме того, осуществляют селективный транспорт. Пассивный транспорт веществ с помощью переносчиков называется облегченной диффузией.
Иногда требуется перенести вещество из области с меньшим значением электрохимического потенциала в область с большим его значением. Этот процесс не может протекать самопроизвольно и требует затрат энергии. Такой вид транспорта называется активным. Например, в сторону увеличения электрохимического потенциала осуществляется трансмембранный перенос натрия. Если энергия, необходимая для осуществления активного транспорта, берется за счет гидролиза АТФ или окислительно-восстановительных реакций, то такой транспорт называется первично-активным; если – за счет градиента концентраций других ионов, то – вторично-активным или сопряженным.
Через мембрану могут переноситься не только отдельные молекулы, но и твердые тела (фагоцитоз), растворы (пиноцитоз). Если вещество транспортируется внутрь клетки, то такой вид транспорта называется эндоцитозом, если наружу, то – экзоцитозом. В первом случае на наружной стороне мембраны образуется впячивание, которое постепенно превращается в пузырек. Пузырек отрывается от мембраны внутри клетки. Такой пузырек содержит в себе транспортируемое вещество, окруженное билипидной оболочкой (везикулой). В дальнейшем везикула сливается с какой-нибудь клеточной органеллой и выпускает в неё своё содержимое. В случае экзоцитоза процесс происходит в обратной последовательности: везикула подходит к мембране с внутренней стороны клетки, сливается с ней и выбрасывает своё содержимое в межклеточное пространство.
1. Пассивный транспорт - перенос молекул и ионов через мембрану, который осуществляется в направлении меньшей их концентрации. Пассивный транспорт не связан с затратой химической энергии. Он стремится выровнять концентрации частиц по разные стороны от мембраны, т.е. свести к нулю величины их градиентов. Если бы в клетках существовал только пассивный транспорт, то значения физической величины внутри и вне клетки сравнялись бы, но этого не происходит.
Различают несколько типов пассивного транспорта (рис. 11.7).
• Простая диффузия через липидный слой. Она подчиняется уравнению Нернста-Планка. В живой клетке такая диффузия обеспечивает прохождение кислорода и углекислого газа, ряда лекарственных веществ. Однако простая диффузия протекает достаточно медленно и не может снабдить клетку в нужном количестве питательными веществами.
• Транспорт через каналы (поры). Канал - участок мембраны, включающий белковые молекулы и липиды, который образует в мембране проход. Этот проход допускает проникновение через мембрану молекул воды, крупных ионов. Наличие каналов увеличивает проницаемость Р. Проницаемость Р зависит от числа каналов и от их радиуса. Каналы могут проявлять селективность
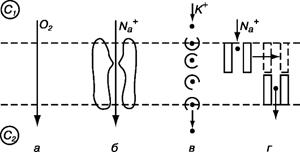 Рис. 11.7. Виды пассивного транспорта: простая диффузия (а), транспорт через каналы (б), облегченная диффузия (в), эстафетная передача (г)
Рис. 11.7. Виды пассивного транспорта: простая диффузия (а), транспорт через каналы (б), облегченная диффузия (в), эстафетная передача (г)
по отношению к разным ионам, это выражается в различной проницаемости для разных ионов.
• Облегченная диффузия - перенос ионов специальными молекулами-переносчиками за счет диффузии переносчика вместе с веществом. Наиболее подробно это явление изучено для случая переноса ионов некоторыми антибиотиками, например валиномицином. Установлено, что валиномицин резко повышает проницаемость мембраны для ионов К+ благодаря специфике своей структуры. В нем формируется полость, в которую точно и прочно вписывается ион К+ (ион Na+ слишком велик для отверстия в молекуле валиномицина). Молекула валиномицина, «захватив» ион К+, образует растворимый в липидах комплекс и проходит через мембрану, затем ион К+ остается, а переносчик уходит обратно.
• Эстафетная передача. В этом случае молекулы-переносчики образуют временную цепочку поперек мембраны и передают друг другу диффундирующую молекулу.
2. Активный транспорт - перенос молекул и ионов, который происходит с затратой химической энергии в направлении от меньших значений величин к большим. При этом нейтральные молекулы переносятся в область большей концентрации, а ионы переносятся против сил, действующих на них со стороны электрического поля. Таким образом, активным транспортом осуществляется перенос веществ в направлении, противоположном транспорту, который должен был бы происходить под действием градиентов (прежде всего концентрационного и электрического). Энергия получается за счет гидролиза молекул особого химического соединения - аденозинтрифосфорной кислоты (АТФ). Экспериментально установлено, что энергии распада одной молекулы АТФ достаточно для выведения наружу трех ионов натрия и введения внутрь клетки двух ионов калия. Одна из схем активного транспорта представлена системой на рис. 11.8.
Захватив одним активным центром ион калия из наружной среды, а другим ион натрия - из внутренней, система, потребляя АТФ, поворачивается внутри мембраны на 180°. Ион натрия оказывается вне клетки и там отделяется, а ион калия попадает внутрь и тоже освобождается, после чего молекула белка принимает исходное положение, и все начинается сначала.
За счет активного транспорта клетка поддерживает внутри себя высокую концентрацию калия и низкую концентрацию натрия. При
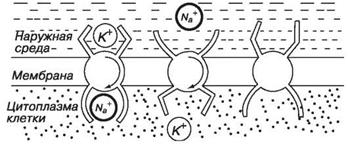 Рис. 11.8. Схема активного транспорта
Рис. 11.8. Схема активного транспорта
этом ионы могут перемещаться против градиента их концентрации (аналогия с газом: перекачивание газа из сосуда с низким давлением в сосуд с высоким давлением).
Активный транспорт обеспечивает механизм селективной проницаемости клеточных мембран. Активный транспорт - важнейшая особенность жизненных процессов.
Основные понятия и формулы
 Окончание таблицы
Окончание таблицы

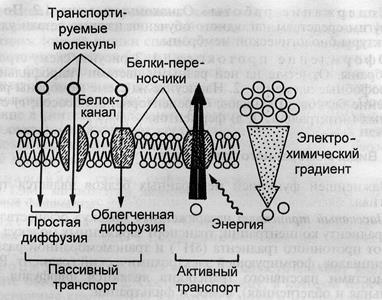
Активный: вещества переносятся через мембрану против концентрационного, электрического и других видов градиентов, на это тратится энергия клеточного метаболизма. Первичный активный транспорт и вторичный активный транспорт.
Пассивный: вещество без затрат энергии клеточного метаболизма переносится через мембрану в направлении градиента. В его основе диффузия и осмос.
- Диффузия вещества в однородной среде
Рассмотрим ситуацию, когда в однородную жидкую (газообразную) среду введено некоторое количество инородного вещества. Вначале распределение этого вещества по объему жидкости будет неравномерным. Однако с течением времени вследствие явлений переноса концентрации этого вещества в различных областях жидкости будут выравниваться.
Диффузия в однородной среде - явление самопроизвольного переноса массы вещества из области с большей концентрацией в область с меньшей концентрацией. Такой перенос называется пассивным транспортом.
Количественно диффузия описывается специальными параметрами.
1. Поток вещества через некоторую поверхность.
В пространстве, заполненном частицами диффундирующего вещества, выделим некоторое направление ОХ, вдоль которого изменяется концентрация частиц, и небольшой элемент поверхности, перпендикулярный этому направлению.
Потоком вещества (Ф) через элемент поверхности, который перпендикулярен направлению диффузии, называется количество этого вещества, переносимого через данный элемент за единицу времени.
Количество переносимого вещества можно измерять в килограммах или молях (ν). В зависимости от этого поток определяется формулами:
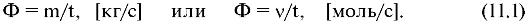 Очевидно, что поток пропорционален площади S выделенного элемента. Кроме того, можно показать, что поток пропорционален градиенту концентрации (dc/dx) диффундирующего вещества в направлении ОХ. Поэтому имеет место следующая формула для расчета потока:
Очевидно, что поток пропорционален площади S выделенного элемента. Кроме того, можно показать, что поток пропорционален градиенту концентрации (dc/dx) диффундирующего вещества в направлении ОХ. Поэтому имеет место следующая формула для расчета потока:
 Коэффициент пропорциональности D называется коэффициентом диффузии. Знак «-» означает, что поток направлен в сторону убывания концентрации вещества (т.е. перенос происходит из области с большей концентрацией в область с меньшей концентрацией).
Коэффициент пропорциональности D называется коэффициентом диффузии. Знак «-» означает, что поток направлен в сторону убывания концентрации вещества (т.е. перенос происходит из области с большей концентрацией в область с меньшей концентрацией).
В формуле (11.2) можно использовать как массовую плотность, так и молярную плотность. Этим и определяется единица измерения потока (кг/с или моль/с). Независимо от единиц плотности размерность коэффициента диффузии D - [м2/с].
Коэффициент D зависит от свойств жидкости, свойств диффундирующих частиц, температуры. Его численное значение выражается формулой: D = σ2/(3τ), где σ - среднее перемещение молекул (среднее расстояние между молекулами), τ - среднее время «оседлой жизни» молекулы.
2. Плотность потока вещества.
Плотностью потока вещества (J) называется отношение потока вещества (Ф) через элемент поверхности к площади этого элемента (S):
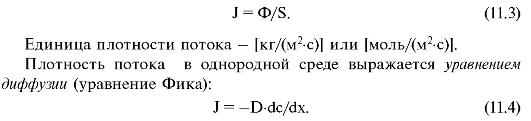 Знак «-» показывает, что суммарная плотность потока вещества при диффузии направлена в сторону, противоположную градиенту концентрации (dc/dx).
Знак «-» показывает, что суммарная плотность потока вещества при диффузии направлена в сторону, противоположную градиенту концентрации (dc/dx).
Уравнение Фика описывает диффузию в однородной среде. Модифицируем его для случая диффузии через мембрану. Обратим внимание на следующий известный факт: на границе раздела двух сред (например, воды и масла) обязательно имеет место скачкообразное изменение концентрации частиц диффундирующего вещества. Например, если в сосуд, в котором поверх воды налито масло, бросить соль, то ее концентрации в этих средах будут различны.
Коэффициент распределения вещества (К) - величина, равная отношению концентраций частиц в граничащих средах:
 Коэффициент распределения вещества - величина безразмерная.
Коэффициент распределения вещества - величина безразмерная.
Рассмотрим диффузию незаряженных частиц (молекул или атомов) через поверхности мембраны. На рисунке 11.6 обозначены:
сi - концентрация частиц внутри клетки (i - от in);
смi - концентрация частиц в мембране у ее внутренней поверхности;
смо - концентрация частиц в мембране у ее внешней поверхности;
со - концентрация частиц вне клетки (о - от out).
Коэффициент распределения вещества между мембраной и окружающей средой равен коэффициенту распределения вещества между мембраной и клеткой:
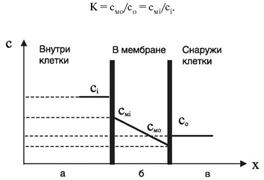 Рис. 11.6. Распределение концентрации частиц, проходящих через мембрану
Рис. 11.6. Распределение концентрации частиц, проходящих через мембрану
Отсюда получаются выражения для концентрации частиц внутри мембраны:
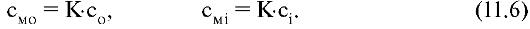 Величины со и с можно измерить.
Величины со и с можно измерить.
Учитывая малую толщину мембраны (L), можно считать, что концентрация молекул диффундирующего вещества изменяется в ней линейно. Поэтому градиент концентрации диффундирующего вещества постоянен:
 Коэффициент проницаемости Р характеризует способность мембраны пропускать те или иные вещества.
Коэффициент проницаемости Р характеризует способность мембраны пропускать те или иные вещества.
Осмос
1748 г. – открытие осмоса. Офицально считается, что открыл Жан-Антуан Молле. Особые свойства мочевого пузыря лягушки. установил, что эта мембрана обладает особым свойством: если по одну сторону чистая вода, по другую растворенные вещества (растворы сахаров). В этих условиях вода начинает активно проникать через мембрану мочевого пузыря в раствор.
Осмос заключается в переходе молекул воды через мембрану по направлениям ее концентрационных градиентов. Наступает равновесие (динамическое) определяется фактором осмотического давления (направление слева направо).
Гидростатическое давление раствора в правом отсеке, когда эти два давления уравновесили друг друга, то мы получим равновесие. Вывод: для того, чтобы измерить осмотическое давление раствора нужно измерить гидростатическое давление во втором отсеке.
В 1877 г. Пфейффер определил количественный показатель осмоса с помощью осмометра (имеет полупроницаемую мембрану – из осадочного ферроцианида Сu). Пфейффер сделал заключение – осмотическое давление пропорционально концентрации растворенного вещества.
Вант-Гофер: в термодинамическом отношении молекулы воды ведут себя подобно молекулам газа.
π=RTS или π=RTη/V,
π – осмотическое давление,
RTη – количество молей вещества,
R – газовая постоянная,
Т – абсолютная температура,
С – концентрация.
Это выражение справедливо лишь для разбавленных растворов.
Осмотичность:
два раствора, в которых создается одинаковое осмотическое давление по обе стороны мембраны проницаемой только для воды называются изоосмотическими, растворы содержат в единице объема одинаковое число растворенных молекул. Если один из растворов имеет осмотическое давление по отношению к другому, то первый раствор называется гиперосмотически, второй – гипоосмотическим.
Тоничность:
определяется по реакции клеток и тканей на их погружение в раствор; если при погружении в раствор ткань не набухает, не сморщивается, такой раствор называют изотоническим по отношению к ткани. Если при погружении ткань набухает – раствор гипотонический, если ткань сморщивается – раствор гипертонический.
- Пассивный перенос вещества через мембрану
При наличии градиента концентраций вещества совершается диффузия или пассивный транспорт указанного вещества из области с большей концентрацией в область с меньшей концентрацией. Это явление происходит самопроизвольно (без затрат энергии) до тех пор, пока концентрации не выровняются, и суммарный поток вещества не обратится в ноль (в случае живых клеток такое выравнивание может и не наступить, если вещества непрерывно синтезируются или, наоборот, затрачиваются в ходе химической реакции).
Различают несколько типов пассивного переноса веществ через мембраны: простая диффузия, перенос через каналы и транспорт с помощью переносчиков за счет диффузии переносчика вместе с веществом в мембране (подвижный переносчик) или эстафетной передачи вещества от одной молекулы переносчика к другой (молекулы переносчика образуют временную цепочку поперек мембраны). Все механизмы переноса веществ делят на две группы:
Такие, при которых каждая молекула переносится независимо от других и эффекты концентрационного насыщения отсутствуют;
Такие, в которых перенос осуществляется после связывания транспортируемой молекулы переносчиком; по мере заполнения свободных переносчиков наблюдается эффект концентрационного насыщения скорости переноса.
Пассивный перенос вещества вдоль оси х описывается уравнением Фика: Φ = –Ddc/dx, где Φ – поток вещества; D – коэффициент диффузии; dc/dx – градиент концентрации c в направлении x. Знак «-«означает, что поток направлен в сторону меньших значений концентрации, то есть приводит к уменьшению величины концентрационного градиента. Для расчетного описания переноса веществ через биологическую мембрану пользуются з а к о н о м Ф и к а д л я п а с с и в н о г о т р а н с п о р т а в е щ е с т в ч е р е з м е м б р а н у: Φ = –DK/l •(cвн-cвв) = –P(cвн-cвв), где Φ – плотность потока; D – коэффициент диффузии; К – коэффициент распределения вещества между мембраной и окружающей водной фазой; l – толщина мембраны; cвв – концентрация частиц внутри клетки; свн – концентрация частиц снаружи клетки; P – коэффициент проницаемости (см. тему 2). Иными словами, величина потока и скорость транспорта веществ через мембрану прямо пропорциональны коэффициенту распределения, который количественно отражает степень липофильности вещества. Чем больше значение коэффициента распределения, тем лучше вещество растворяется в мембране и с тем большей скоростью переносится через неё. Если рассматривать пассивный переход с позиций превращения энергии, то поток, проходящий через биологическую мембрану, равен: Φ = – uc (dG/dx), где u = D/RT - коэффициент пропорциональности, который зависит от скорости диффузии молекул и называется подвижностью. Таким образом, поток пропорционален концентрации вещества и градиенту термодинамического потенциала в направлении тока.
- Перенос заряженных частиц через мембрану
Пассивный транспорт ионов.
1.
В том случае, когда изменяется концентрация вещества и электрический потенциал, уравнения для потоков незаряженных и заряженных частиц сходны: Φ = – uc (dG/dx).
2.
Если изменяется еще и стандартный электрохимический потенциал, то уравнение для потока вещества описывается уравнением Теорелла: Φ = – cu • (dμ/dx), где μ – электрохимический потенциал, который зависит от природы вещества и природы растворителя.
3.
Если во всей рассматриваемой области диффузии отсутствуют химические превращения вещества и растворитель одинаков, то в этом случае уравнение Теорелла сводится к электродиффузному уравнению Нернста-Планка: Φ = –uRT (dc/dx) –cuz• Fdφ/dx. Это уравнениеописывает пассивный перенос частиц в условиях существования градиентов концентрации вещества и электрического потенциала в растворе или в однородной незаряженной мембране.
4.
Если известны концентрации ионов по обе стороны мембраны, трансмембранная разность потенциалов и коэффициент проницаемости, то величину пассивного потока ионов описывают с помощью уравнения Гольдмана: Φ ={zFΔφΡ/RT} • {[c2exp(zFΔφ/RT) -c1]/[1-exp(zFΔφ/RT]}.
5.
Если потоки направлены противоположно, то их отношение описывается уравнением Уссинга-Теорелла: Φ2/Φ1 = c1/c2exp[- zFΔφ/RT].
Вывод: пассивный транспорт ионов может происходить только в случае наличия на мембране градиентов концентрации или электрического потенциала и описывается в общем случае уравнениями Нернста-Планка, Гольдмана или Уссинга-Теорелла. Невыполнение этих уравнений свидетельствует о том, что перенос ионов осуществляется активно или через мембранные каналы, где невозможно существование независимых ионных потоков.
Уравнение Нернста. В живом организме по обе стороны любой биологической мембраны находятся ионные растворы, причем концентрации одного и того же иона по обе стороны часто отличаются. Одной из причин установления и поддержания этих градиентов концентраций является различная проницаемость мембраны для тех или иных ионов. В этом случае градиент концентрации одного вещества может поддерживаться за счет наличия градиента концентрации другого вещества и/или разности потенциалов по обе стороны мембраны (так называемой трансмембранной разности потенциалов). Условием установления равновесия между двумя растворами, разделенными полупроницаемой мембраной, с различными концентрациями одинаковых ионов является равенство электрохимических потенциалов по одну и другую сторону мембраны. Значение разности электрических потенциалов, которая устанавливается на мембране при наличии градиента концентраций, рассчитывается с помощью уравнения Нернста: Δφ = φi - φ0 = (RT/zF)•lg (c0/ci).
Уравнение Фика описывает как пассивный транспорт незаряженных частиц, так и пассивный транспорт заряженных частиц в отсутствии электрического поля. Рассмотрим теперь транспорт ионов с учетом электрического поля внутри мембраны.
На отдельный ион в электрическом поле действует сила f0 = qE, где Е - напряженность электрического поля, а q = Ze - заряд иона (Z - валентность иона). Напряженность поля выражается через градиент
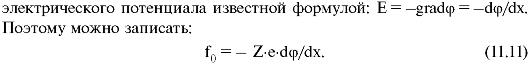 Уравнению 11.11 можно придать более удобный вид, умножив обе его части на постоянную Авогадро (NA):
Уравнению 11.11 можно придать более удобный вид, умножив обе его части на постоянную Авогадро (NA):  Это уравнение Нернста-Планка (электродиффузное уравнение). Оно устанавливает зависимость плотности диффузного потока ионов от концентрации ионов и от градиента потенциала (dφ/dx).
Это уравнение Нернста-Планка (электродиффузное уравнение). Оно устанавливает зависимость плотности диффузного потока ионов от концентрации ионов и от градиента потенциала (dφ/dx).
Для нейтральных частиц (Z = 0) уравнение Нернста-Планка переходит в уравнение Фика.
Равновесие Доннана. В клетке кроме малых ионов находятся еще и заряженные макромолекулы (белки, нуклеиновые кислоты). Как правило, эти макромолекулы заряжены отрицательно. Мембрана проницаема для малых ионов и непроницаема для макромолекул. Для каждого из растворов, находящихся по обе стороны мембраны, необходимо выполнение условия электронейтральности, то есть сумма положительных и отрицательных зарядов всех ионов должна равняться нулю. Для компенсации внутри клетки отрицательных зарядов макромолекул часть отрицательных малых ионов выходит из клетки, а часть положительных зарядов проходит в клетку из межклеточной среды. То есть на мембране возникает так называемая доннановская разность потенциалов, которая составляет φi - φ0 = -RT/F•[R-]i /2c.
Ионный транспорт через каналы. Механизм обычной электродиффузии в живой клетке обеспечивает проницаемость мембран для кислорода и углекислого газа. Этот процесс происходит слишком медленно и плохо контролируется, поэтому клетка не может его использовать для переноса питательных веществ и необходимых для жизнедеятельности ионов. Так, из каждых 1061 ионов, находящихся в водном растворе, только один находится в липидной фазе мембраны. Скорость переноса ионов значительно возрастает, если в мембране существуют ионные каналы. Наиболее распространены каналы для ионов калия, натрия, кальция. Ионные мембранные каналы образованы интегральными белками. Такой канал может быть либо открыт, либо закрыт для транспорта ионов. Эти два состояния реализуются изменением конформации каналообразующих белков, что может быть вызвано изменением условий в клетке и во внеклеточной среде, например, изменением мембранного потенциала.
Так как обычно с помощью каналов транспортируются гидрофильные вещества, то во внутренней полости их имеется большое число гидрофильных химических групп. Каждый канал неоднороден по строению: вдоль его внутренней полости располагаются различные химические группы, сродство ионов к которым неодинаково. Канал может иметь один или несколько ионных центров связывания. Эти центры представляют собой заряженные группы. Когда ион попадает в канал, он связывается с этими группами и таким образом попадает в потенциальную яму. Для того чтобы попасть в другую потенциальную яму, иону требуется преодолеть некоторый потенциальный барьер. Таких барьеров вдоль длины канала может быть несколько, причем высота их, обычно, неодинакова и может изменяться в зависимости от наличия или отсутствия ионов в канале, или изменения трансмембранной разности потенциалов. Так как в канале находятся заряженные группы, то изменение мембранного потенциала является нелинейным. Кроме этого, наличие заряженных групп может привести к непостоянству значений коэффициента распределения по длине канала.
Многие мембранные каналы настолько узки, что ионы не могут в них двигаться в различных направлениях независимо друг от друга: если в какой-либо потенциальной яме уже есть ион, то другой не может в неё попасть. Это относится, например, к калиевым каналам. Встречные потоки ионов натрия через натриевые каналы можно считать независимыми, но если через эти каналы движутся ионы калия, то принцип независимости уже не выполняется. Для многих каналов, в зависимости от их строения, при высоких концентрациях ионов в окружающей среде наблюдается а) э ф ф е к т н а с ы щ е н и я: при увеличении концентрации ионов скорость их переноса повышается, но только до определенного уровня, выше которого скорость не изменяется; б) б л о к и р о в к а: при концентрациях ионов выше некоторого значения скорость переноса начинает снижаться.
При переходе иона из окружающей среды в канал свободная энергия уменьшается. При этом величина потенциального барьера ниже, чем при переходе иона непосредственно через липидный бислой мембраны.
Ионные каналы обладают селективностью, то есть имеют различные величины проводимости для различных ионов. Каждый канал транспортирует преимущественно ионы одного вида, например, натриевый канал – ионы натрия, калиевый – ионы калия. Через катионные каналы не способны проходить анионы и наоборот. Но для ионов того же знака, что и основной ион, канал уже не обладает такой абсолютной селективностью. Проводимость канала для других ионов своего знака ниже, чем для собственного, но отлична от нуля. Например, через специфические натриевые каналы способны транспортироваться также и ионы калия, но в 20 раз медленнее, чем ионы натрия.
Согласно современным представлениям, селективность канала, прежде всего, определяется радиусом транспортируемого иона. Ионы, радиус которых превышает радиус канала, не способны проходить через канал вследствие такого стерического несоответствия. Ионы, обладающие меньшим радиусом, по сравнению с радиусом канала, также не всегда могут пройти через канал. Чем меньше радиус иона по сравнению с радиусом канала, тем меньше вероятность прохождения иона через этот канал. Этот эффект объясняется тем, что ионы в водном растворе всегда находятся в гидратированном состоянии. Перенос же через канал требует сброса нескольких гидратных оболочек, в результате которого ион остается лишь в одной гидратной «одежке». Сброшенная гидратная оболочка заменяется другими полярными соединениями, в роли которых выступают фиксированные дипольные группы канала. Размер иона должен быть таковым, чтобы он мог взаимодействовать с оптимальным числом этих полярных групп. Например, ион калия в своем селективном канале взаимодействует со всеми пятью атомами кислорода, а меньший по размеру ион натрия может связаться в калиевом канале только с двумя атомами кислорода. Следовательно, прочность связи канала с ионом натрия будет минимальна. А если связи иона с дипольными группами нет совсем, то и пройти он через этот канал не может. Следовательно, лучше всего проходит через канал тот ион, который имеет радиус (вместе с одинарной гидратной «одежкой») равный радиусу канала.
Селективность каналов определяется не только зависимостью проводимости канала от радиуса транспортируемого иона, но и химическим строением иона. Например, через натриевые каналы переносятся ионы гидроксиламмония и не переносятся ионы метиламмония, хотя их радиусы равны радиусу канала. Это объясняется тем, что гидроксильная группа гидроксиламмония способна образовывать водородные связи с атомами кислорода, выстилающими внутренний диаметр канала, а метильная группа метиламмония с ними не взаимодействует.
Каналы могут блокироваться некоторыми соединениями, которые связываются с селективными фильтрами или с входом в канал, закупоривая его. Так как молекула-блокатор должна стерически соответствовать участку блокируемого канала, то для каналов определённого вида существуют специфические блокаторы. Например, молекулы тетродоксина блокируют натриевые каналы, а молекулы тетраэтиламмония – калиевые.
- Активный транспорт
В организме часто наблюдается транспорт нейтральных частиц или ионов, в процессе которого электрохимический потенциал возрастает. Например, концентрация ионов калия в клетке намного выше, чем в межклеточной среде, а натрия, наоборот, - ниже. Высокий градиент концентрации калия поддерживается градиентом электрического потенциала, направленным в противоположную сторону, и подчиняется условию равновесия, определяемым уравнением Нернста. Направление градиента электрического потенциала таково, что ионам натрия также гораздо выгоднее находиться в клетке, чем во внеклеточной среде. Самопроизвольный вынос из клетки ионов натрия невозможен и осуществляется только за счет притока энергии. Известны системы активного транспорта дл ионов калия, натрия, кальция и водорода.
Поддержание градиента концентраций ионов натрия и калия обеспечивается работой натрий-калиевого насоса. Он представляет собой мембранные белки, которые работают как ферменты аденозинтрифосфатазы (АТФазы). Задачей этих ферментов является расщепление АТФ на АДФ и неорганический фосфат. Процесс распада сопровождается выделением энергии, которая расходуется на транспорт ионов в сторону увеличения электрохимического потенциала. Расщепление АТФ стимулируется ионами натрия и калия и зависит от наличия магния. Активный транспорт возможен только за счет сопряжения транспорта какого-либо вещества с реакцией гидролиза АТФ.
Механизм сопряжения окончательно не выяснен. Вероятнее всего, энергия АТФ расходуется на изменение конформации транспортного белка, что изменяет его сродство (константу связывания) к тем или иным ионам. Транспорт всегда осуществляется в ту сторону, где сродство ниже. В клетке константа связывания переносчика с Na+ значительно выше, чем с K+. Поэтому ионы натрия в клетке связываются с белком и транспортируются во внеклеточную среду. По другую сторону мембраны конформация белка меняется таким образом, что константа связывания с Na+ уменьшается, а с K+ – увеличивается. Структура ионсвязывающего участка белка в этом случае такова, что к нему могут присоединяться уже не три, а два иона калия, которые и переносятся в клетку.
Согласно современным представлениям, процесс активного транспорта Na+ и K+ происходит в следующие семь этапов.
1.
В присутствии Mg2+ на внутренней стороне мембраны образуется комплекс фермента АТФазы с АТФ.
2.
Присоединение АТФ изменяет конформацию фермента таким образом, что к образовавшемуся комплексу присоединяются три иона натрия.
3.
Происходит фосфорилирование Na+,K+ - АТФазы и отщепление АДФ.
4.
Ионсвязывающий центр фермента перемещается относительно толщины мембраны, в результате чего ион натрия оказывается на внешней стороне клетки.
5.
Снаружи клетки вследствие уменьшения сродства фермента к ионам натрия и повышения сродства к калию происходит обмен этими ионами.
6.
После отщепления фосфата фермент с присоединенными ионами калия снова изменяет положение относительно мембраны.
7.
Ионы калия и неорганический фосфат высвобождаются в цитоплазму, и фермент возвращается в исходное состояние.
Таким образом, энергии, выделяющейся при гидролизе одной молекулы АТФ достаточно, чтобы вынести из клетки три иона натрия и внести два иона калия. Na+,K+ - насос способствует не только повышению градиентов концентраций ионов, но и возрастанию градиента электрического потенциала, то есть является электрогенным, так как сумма вносимых зарядов неравна сумме выносимых. Межклеточная среда приобретает «более положительный» заряд по сравнению с клеткой за счет выноса одного «лишнего» положительного иона.
В мембранах саркоплазматического ретикулума мышечных клеток и цитоплазматических мембранах кардиомиоцитов существует Ca2+ - насос, работа которого во многом сходна с механизмом переноса ионов Na+,K+ - насосом. За один цикл, в процессе которого расходуется одна молекула АТФ, переносится два иона кальция.
Активный транспорт протонов может осуществляться как с помощью подвижных переносчиков, так и через мембранные каналы. Протонные каналы представляют собой интегральные белки, образующие внутреннюю пору, где содержатся участки, к которым могут присоединяться протоны. Энергия АТФ расходуется на изменение конформации белковых молекул, вследствие чего сродство одних участков связывания к протонам понижается, а других – увеличивается, что заставляет протон перескочить на другой участок канала, сродство которого к протону на данный момент выше. Путем таких перескоков с одного участка связывания на другой ион водорода пересекает мембрану.
Перенос Н+ против градиентов их концентраций осуществляется не только за счет энергии, выделяющейся при гидролизе АТФ, но и за счет энергии фотонов. Этот способ используется галофильными бактериями, которые на свету выкачивают протоны из клетки, а энергию создавшегося градиента концентраций используют для синтеза АТФ.
Вторично-активный транспорт. Транспорт, источником энергии которого служит не непосредственно АТФ или энергия окислительно-восстановительных реакций, а градиент концентраций других веществ, называется вторично-активным, или сопряженным транспортом. Существует три вида вторично-активного транспорта ионов: унипорт, симпорт и антипорт.
1.
В случае унипорта за счет существования на мембране градиента электрического потенциала осуществляется однонаправленный транспорт заряженных частиц в сторону меньшего значения потенциала. Например, митохондрии в процессе своего функционирования активно выкачивают протоны из матрикса в цитоплазму, в результате чего их внутренняя область оказывается заряженной более отрицательно, чем внешняя. В нормальных условиях созданный градиент электрохимического потенциала используется для синтеза АТФ.
2.
В процессе антипорта осуществляется транспорт одинаково заряженных ионов двух типов в разные стороны.
3.
По механизму симпорта осуществляется транспорт противоположно заряженных ионов в одну сторону. При этом транспорт одного из ионов осуществляется по градиенту концентраций, а транспорт второго - по градиенту электрического потенциала, создаваемого транспортом первого иона.
Рис. 3. Схема вторично-активного транспорта веществ.
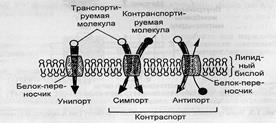
Одним из наиболее изученных случаев вторично-активного транспорта незаряженных молекул является всасывание глюкозы в стенках кишечника. Концентрация глюкозы в энтероцитах выше, чем в просвете кишечника, поэтому пассивное её всасывание невозможно. Клетки кишечника активно выкачивают натрий из энтероцитов в серозную область кишечника, в результате чего концентрация Na+ в клетке снижается по сравнению с его концентрацией снаружи. Благодаря этому становится возможным пассивный транспорт натрия из просвета кишечника в энтероцит. Однако простая диффузия ионов через бислой мало вероятна и натрий может пересечь мембрану только с помощью переносчика. При этом переносчик связывается не только с натрием, но и с молекулой глюкозы. Образовавшийся комплекс пассивно по градиенту концентраций ионов натрия и электрического потенциала переносится внутрь клетки. Таким образом, транспорт глюкозы осуществляется против градиента концентрации, но не за счет энергии АТФ, а за счет существования градиента концентрации другого вещества, в данном случае – ионов натрия. Подобные системы транспорта существуют и для многих других веществ, например, углеводов и аминокислот, что очень важно, так как для них отсутствуют специфические насосы.
- Образование мембранных потенциалов
В ходе эволюции у железистого эпителия, мышечной и нервной тканей появилось свойство возбудимости – способность реагировать на воздействие окружающей среды возбуждением. Внешним проявлением возбуждения является возникновение биоэлектрического потенциала.
Все ткани организма могут находиться в двух состояниях:
1. состоянии относительного физиологического покоя;
2. состоянии активности.
Наблюдается при раздражении ткани. Существует 2 вида активного состояния тканей: возбуждение и торможение. Возбуждение – это активный процесс, представляющий собой ответную реакцию ткани на раздражение и характеризующийся повышением функций ткани. Возбуждение характеризуется двумя группами признаков: неспецифическими и специфическими.
Неспецифические признаки возникают у всех возбудимых тканей вне зависимости от их строения:
1. изменение проницаемости клеточных мембран
2. изменение заряда клеточных мембран,
3. повышение потребления кислорода
4. повышение температуры
5. усиление обменных процессов
Специфические признаки различаются у различных тканей:
1. мышечная ткань – сокращение
2. железистая ткань – выделение секрета
3. нервная ткань – генерация нервного импульса.
Процесс возбуждения связан с наличием в мембране электрически (для ионов кальция и хлора) и химически (для ионов натрия и калия) управляемых каналов, которые могут открываться в ответ на соответствующее раздражение клетки.
Мембранный потенциал, разность электрических потенциалов между растворами электролитов a и b, разделенных проницаемой мембраной m: Dabj = ja - jb. В частном случае, когда мембрана проницаема только для определенногоиона Вzв (zB - зарядовое число), общего для растворов электролитов a и b, мембранный потенциал (иногда его называют потенциалом Нернста) рассчитывают по формуле:

где F -число Фарадея, R -газовая постоянная, Т -абсолютная температура, aB b, aB a - активности ионов. В растворах b и a, DabjB-стандартный потенциал распределения иона В, равный
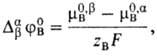
где m0,bB, m0,aB-стандартные хим. потенциалы иона В в растворах b и a соответственно. В такой системе мембранный потенциал не зависит от толщины мембраны и ее структуры, механизма переноса иона и его подвижности.
В общем случае для мембран, проницаемых для одних сортов ионов и не проницаемых для других, при расчете мембранный потенциал требуется введение определенных приближений в зависимости от толщины мембраны, ее состава и строения, а также от механизма переноса. В случае мембран макроскопического размера полный мембранный потенциал слагается из трех компонентов: двух граничных потенциалов, локализованных в двойных электрических слоях на границе мембрана-раствор электролитов, и внутримембранного, локализованного в электронейтральном объеме мембраны. Кроме того, при пропускании электрического тока через мембрану внутри нее возникает падение напряжения. Обычно считается, что переход ионов через межфазную границу происходит быстро, так что их распределение равновесно; затруднен только перенос ионов через объем мембраны. Для системы, в которой в фазе a имеется бинарный электролит В+ А-, присутствующий и в мембране, а мембрана содержит ион R с зарядовым числом zR, не проникающий через межфазные границы, граничный потенциал определяется формулой Доннана и называется доннановским:
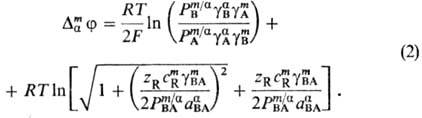
Здесь c m R - концентрация ионов R в мембране, g i - и Рi -коэффициент активности и распределения ионов соответственно, определяемые соотношением
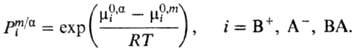
Средняя активность электролита 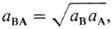 средний коэф. активности
средний коэф. активности 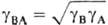 , коэффициент распределения электролита
, коэффициент распределения электролита 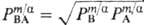
Таким образом, доннановский потенциал (2) состоит из так называемого потенциала распределения и члена, зависящего от концентрации непроникающего иона R; при малом значении c m R он сводится к потенциалу распределения, а при большом - находится из выражения:
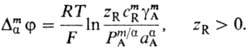
При этом концентрация противоионов А- в мембране перестает зависеть от коэффициента распределения и межфазного потенциала и приближается к предельному значению cm A 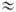 zR cm R, a одноименно заряженные ионы почти полностью вытесняются из мембраны.
zR cm R, a одноименно заряженные ионы почти полностью вытесняются из мембраны.
Граничный потенциал может возникнуть в результате реакций комплексообразования или электронообменной реакции между окислительно-восстановительными парами, содержащимися в растворе и в мембране. Если межфазная граница вообще не проницаема для заряженных частиц, граничный потенциал имеет электростатическую природу и возникает в результате адсорбции зарядов и диполей, а также вследствие заряжения границы от внеш. источника.
Внутримембранный потенциал имеет кинетическую природу и определяется переносом ионов через толщу мембраны. В простейшем случае при диффузионно-миграционном переносе бинарного электролита возникает т. наз. диффузионный потенциал (приближение Планка):
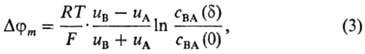
где u А, u B-подвижности ионов в мембране, cm BA(0), cm BA(d)-концентрации электролита в мембране у левой и правой межфазных границ соответственно. В случае биол. и бислойных липидных мембран, толщина которых настолько мала, что в них вообще не реализуется область электронейтральности, для нахождения внутримембранного потенциала используют предположение о постоянстве напряженности электрического поля (приближение Гольдмана):
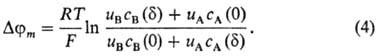
Таким образом, мембранный потенциал в доннановском случае находят из выражения, слагаемые которого определены формулами (2) и (3) или (2) и (4):
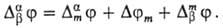
Определение мембранного потенциала представляет интерес для ионометрии, для биологии и медицины в связи с распространенностью мембранных процессов в живых организмах и т.д.
- Потенциал покоя в клетках
Потенциал покоя – относительно стабильная разность электрических потенциалов между наружной и внутренней сторонами клеточной мембраны. Его величина обычно варьирует в пределах от -30 до -90 мВ. Внутренняя сторона мембраны в покое заряжена отрицательно, а наружная – положительно из-за неодинаковых концентраций катионов и анионов внутри и вне клетки.
Внутри- и внеклеточные концентрации ионов (ммоль/л) в мышечных клетках теплокровных животных
| Внутриклеточная концентрация | Внеклеточная концентрация | |
| Na+ | ||
| K+ | ||
| Ca2+ | 10-8 – 10-7 | |
| Cl- | 120 – 130 | |
| HCO3- |
Потенциал покоя - это разность электрических потенциалов между внутренней и наружной сторонами мембраны, когда клетка находится в состоянии физиологического покоя. Его средняя величина составляет -70 мВ (милливольт).
Сущность потенциала покоя - это преобладание на внутренней стороне мембраны отрицательных электрических зарядов в виде анионов и недостаток положительных электрических зарядов в виде катионов, которые сосредотачиваются на её наружной стороне, а не на внутренней.
Внутри клетки - "отрицательность", а снаружи - "положительность".
Такое положение вещей достигается с помощью трёх явлений: (1) поведения мембраны, (2) поведения положительных ионов калия и натрия и (3) соотношения химической и электрической силы.
Date: 2015-09-24; view: 1204; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |