Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Обмен хромо- и нуклеопротеинов
| Раздел 26.1 | Особенности строения и функции хромопротеинов. |
26.1.1. Хромопротеины относятся к сложным белкам. Молекулы хромопротеинов состоят из полипептидных цепей и небелковых компонентов (простетических групп), из которых наиболее распространённым является гем.
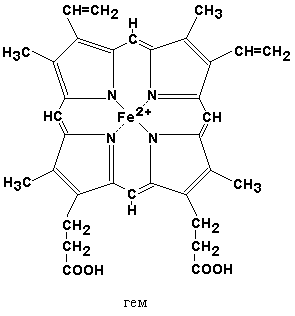 26.1.2. Гем в качестве простетической группы содержат следующие белки:
· Гемоглобин – присутствует в эритроцитах; в его состав входят 4 полипептидные цепи, с каждой из которых связана одна гемовая группа. Этот белок транспортирует О2 и СО2 в крови.
· Миоглобин – присутствует в клетках мышечной ткани; представляет собой одну полипептидную цепь, с которой связана одна гемовая группа. Этот белок запасает кислород в мышцах и отдаёт его при выполнении мышечной работы.
· Цитохромы – белки-ферменты, содержатся в митохондриях клеток, участвуют в переносе электронов на кислород в дыхательной цепи.
· Пероксидаза и каталаза – белки-ферменты, ускоряют расщепление пероксида водорода Н2О2 на Н2О и О2.
26.1.2. Гем в качестве простетической группы содержат следующие белки:
· Гемоглобин – присутствует в эритроцитах; в его состав входят 4 полипептидные цепи, с каждой из которых связана одна гемовая группа. Этот белок транспортирует О2 и СО2 в крови.
· Миоглобин – присутствует в клетках мышечной ткани; представляет собой одну полипептидную цепь, с которой связана одна гемовая группа. Этот белок запасает кислород в мышцах и отдаёт его при выполнении мышечной работы.
· Цитохромы – белки-ферменты, содержатся в митохондриях клеток, участвуют в переносе электронов на кислород в дыхательной цепи.
· Пероксидаза и каталаза – белки-ферменты, ускоряют расщепление пероксида водорода Н2О2 на Н2О и О2.
|
| Раздел 26.2 | Биосинтез гемоглобина. |
26.2.1. Схема биосинтеза гемоглобина представлена на рисунке 26.1. Исходными веществами в этом метаболическом пути являются аминокислота глицин и метаболит цикла Кребса сукцинил-КоА. Синтез происходит в ретикулоцитах (незрелых эритроцитах, содержащих клеточное ядро). Реакции идут в митохондриях и цитоплазме клеток.
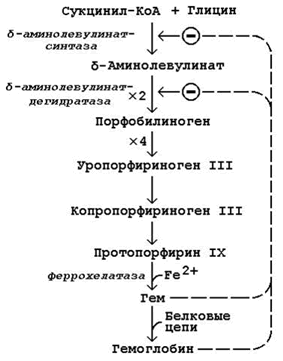 Рисунок 26.1. Биосинтез гемоглобина и его регуляция.
Первая стадия в последовательности реакций, ведущих к синтезу гема, катализируется δ-аминолевулинат-синтазой. Фермент абсолютно специфичен к субстратам; кофакторами фермента являются пиридоксаль-5-фосфат и ионы Mg2+.
Рисунок 26.1. Биосинтез гемоглобина и его регуляция.
Первая стадия в последовательности реакций, ведущих к синтезу гема, катализируется δ-аминолевулинат-синтазой. Фермент абсолютно специфичен к субстратам; кофакторами фермента являются пиридоксаль-5-фосфат и ионы Mg2+.
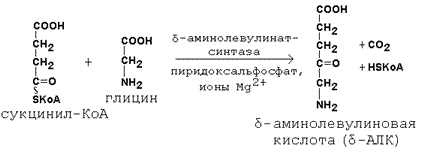 Имеются данные о том, что некоторые лекарственные препараты, а также стероидные гормоны, напротив, индуцируют синтез печёночной δ-аминолевулинат-синтазы.
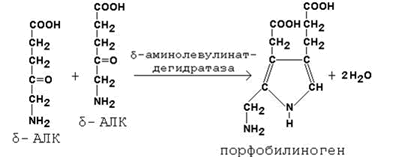
Во второй реакции, катализируемой δ-аминолевулинат-дегидратазой, при конденсации двух молекул δ-аминолевулината образуется порфобилиноген.
Имеются данные о том, что некоторые лекарственные препараты, а также стероидные гормоны, напротив, индуцируют синтез печёночной δ-аминолевулинат-синтазы.
Во второй реакции, катализируемой δ-аминолевулинат-дегидратазой, при конденсации двух молекул δ-аминолевулината образуется порфобилиноген.
 В дальнейшем из четырёх молекул порфобилиногена в результате ряда сложных ферментативных реакций образуется протопорфирин IX – непосредственный предшественник гема. При участии митохондриального фермента феррохелатазы двухвалентное железо включается в уже готовую структуру протопорфирина. Для протекания этой реакции необходимы аскорбиновая кислота и цистеин в качестве восстановителей. Ингибитором феррохелатазы является свинец. На заключительном этапе происходит соединение гема с белковыми цепями, характерными для синтезируемого хромопротеина. Конечные продукты этого биосинтеза (гем, гемоглобин) подавляют начальные реакции по механизму отрицательной обратной связи (рисунок 9).
При врождённых и приобретённых нарушениях биосинтеза гема развиваются заболевания – порфирии.
26.2.2. Порфирии – группа наследственных заболеваний, обусловленных частичным дефицитом одного из ферментов синтеза гема. Снижение образования гема приводит к снятию его ингибирующего эффекта на начальные этапы биосинтеза, результатом чего является избыточное образование порфиринов и их предшественников. Основными симптомами порфирий являются:
· нарушения со стороны центральной нервной системы (т.к. предшественники порфиринов являются нейротоксинами);
· повышенная светочувствительность кожи (порфирины накапливаются в коже, поглощают свет и переходят в возбуждённое состояние, вызывая образование токсичных свободных радикалов);
· анемия(снижение содержания гемоглобина в крови);
· порфиринурия - выведение порфиринов с мочой и калом (моча приобретает красную окраску).
Порфиринурия может также развиваться при отравлениях свинцом.
В дальнейшем из четырёх молекул порфобилиногена в результате ряда сложных ферментативных реакций образуется протопорфирин IX – непосредственный предшественник гема. При участии митохондриального фермента феррохелатазы двухвалентное железо включается в уже готовую структуру протопорфирина. Для протекания этой реакции необходимы аскорбиновая кислота и цистеин в качестве восстановителей. Ингибитором феррохелатазы является свинец. На заключительном этапе происходит соединение гема с белковыми цепями, характерными для синтезируемого хромопротеина. Конечные продукты этого биосинтеза (гем, гемоглобин) подавляют начальные реакции по механизму отрицательной обратной связи (рисунок 9).
При врождённых и приобретённых нарушениях биосинтеза гема развиваются заболевания – порфирии.
26.2.2. Порфирии – группа наследственных заболеваний, обусловленных частичным дефицитом одного из ферментов синтеза гема. Снижение образования гема приводит к снятию его ингибирующего эффекта на начальные этапы биосинтеза, результатом чего является избыточное образование порфиринов и их предшественников. Основными симптомами порфирий являются:
· нарушения со стороны центральной нервной системы (т.к. предшественники порфиринов являются нейротоксинами);
· повышенная светочувствительность кожи (порфирины накапливаются в коже, поглощают свет и переходят в возбуждённое состояние, вызывая образование токсичных свободных радикалов);
· анемия(снижение содержания гемоглобина в крови);
· порфиринурия - выведение порфиринов с мочой и калом (моча приобретает красную окраску).
Порфиринурия может также развиваться при отравлениях свинцом.
|
| Раздел 26.3 | Обмен железа. |
26.3.1.В организме человека содержится 4 – 6 г железа. Из этого количества 65-70% приходится на долю гемоглобина. Значительно меньше Fе находится в составе других гемсодержащих белков (миоглобин, цитохромы), а также металлопротеинов (ферритин, трансферрин). Поэтому обмен железа в организме определяется прежде всего синтезом и распадом гемоглобина эритроцитов. Недостаточное поступление железа в организм проявляется в первую очередь как анемия (железодефицитная). Общая схема обмена железа представлена на рисунке 26.2.
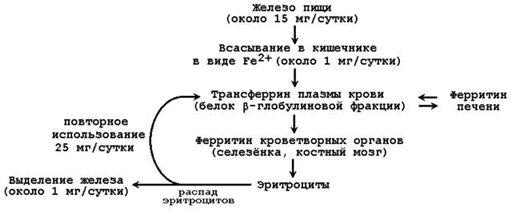 Рисунок 26.2. Обмен железа в организме.
26.3.2.В кишечнике всасывается лишь небольшая часть (около 1/10) имеющегося в пище железа. Транспортной формой железа в крови служит белок плазмы крови трансферрин. Другой белок, участвующий в метаболизме железа – ферритин – служит для депонирования железа, присутствует в большинстве тканей. Железо, освобождающееся при разрушении эритроцитов, может, как правило, повторно использоваться (реутилизироваться) для построения новых молекул хромопротеинов. Однако часть железа теряется организмом, главным образом, с желчью. Эти потери компенсируются поступлением железа с пищей.
Рисунок 26.2. Обмен железа в организме.
26.3.2.В кишечнике всасывается лишь небольшая часть (около 1/10) имеющегося в пище железа. Транспортной формой железа в крови служит белок плазмы крови трансферрин. Другой белок, участвующий в метаболизме железа – ферритин – служит для депонирования железа, присутствует в большинстве тканей. Железо, освобождающееся при разрушении эритроцитов, может, как правило, повторно использоваться (реутилизироваться) для построения новых молекул хромопротеинов. Однако часть железа теряется организмом, главным образом, с желчью. Эти потери компенсируются поступлением железа с пищей.
|
| Раздел 26.4 | Катаболизм гемоглобина | |||
26.4.1.Содержание гемоглобина в крови здоровых людей составляет 130-160 г/л. Гемоглобин крови полностью обновляется в течение 120 дней (продолжительность жизни эритроцита).
Разрушение эритроцитов и начальные этапы катаболизма гема происходят в клетках ретикуло-эндотелиальной системы (РЭС), которые находятся в печени (клетки Купфера), селезёнке, костном мозге. Схема катаболизма гемоглобина в тканях приводится на рисунке 26.3.
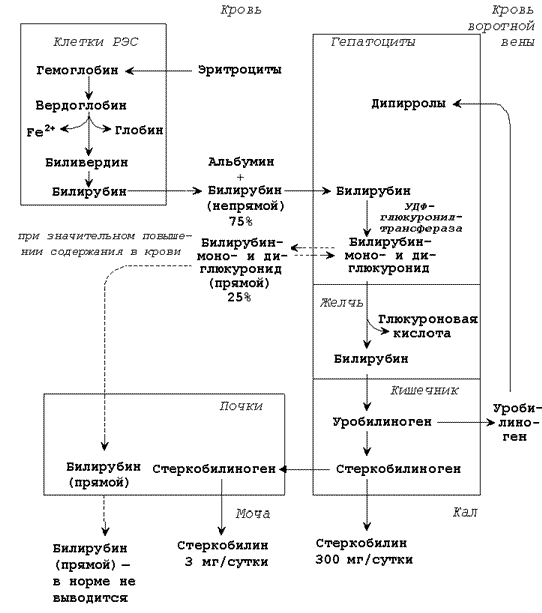 Рисунок 26.3. Схема катаболизма гемоглобина в тканях.
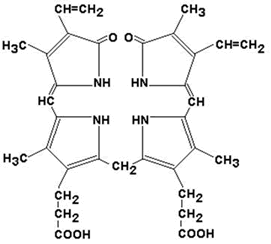
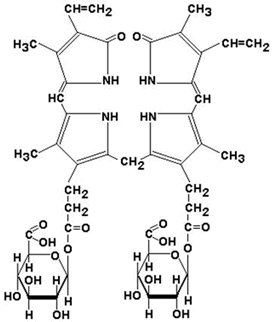
26.4.2. Продукты распада гема называют желчными пигментами, так как все они в разных количествах обнаруживаются в желчи. К желчным пигментам относятся: биливердин (зелёного цвета), билирубин (красно-коричневого цвета), уробилиноген и стеркобилиноген (бесцветные), уробилин и стеркобилин (жёлтого цвета). Далее приводятся формулы билирубина и его диглюкуронида.
Рисунок 26.3. Схема катаболизма гемоглобина в тканях.
26.4.2. Продукты распада гема называют желчными пигментами, так как все они в разных количествах обнаруживаются в желчи. К желчным пигментам относятся: биливердин (зелёного цвета), билирубин (красно-коричневого цвета), уробилиноген и стеркобилиноген (бесцветные), уробилин и стеркобилин (жёлтого цвета). Далее приводятся формулы билирубина и его диглюкуронида.
|
| Раздел 26.5 | Желтухи. |
| 26.5.1.Общее содержание билирубина в крови здорового человека составляет 8 – 20 мкмоль/л, из них 6 – 15 мкмоль/л приходится на непрямой билирубин, 2 – 5 мкмоль/л – на прямой билирубин. Увеличение общего билирубина в крови (более 27 мкмоль/л) приводит к окрашиванию кожи, слизистых оболочек, склеры глаз в жёлтый цвет (желтуха). Определение содержания желчных пигментов в крови используют при выяснении происхождения желтух. Желтуха бывает надпечёночная (гемолитическая), печёночная (паренхиматозная), подпечёночная (обтурационная или механическая). 26.5.2. Надпечёночная (гемолитическая) желтуха вызвана массивным распадом эритроцитов в результате резус-конфликта, попадания в кровь веществ, вызывающих разрушение мембран эритроцитов и некоторых других заболеваниях. При этой форме желтухи в крови повышено содержание непрямого билирубина, в моче повышено содержание стеркобилина, билирубин отсутствует, в кале повышено содержание стеркобилина. 26.5.3. Печёночная (паренхиматозная) желтуха вызвана повреждением клеток печени при инфекциях и интоксикациях. При этой форме желтухи в крови повышено содержание непрямого и прямого билирубина, в моче повышено содержание уробилина, присутствует билирубин, в кале понижено содержание стеркобилина. 26.5.4. Подпечёночная (обтурационная) желтуха вызвана нарушением оттока желчи, например, при закупорке желчевыводящего протока камнем. При этой форме желтухи в крови повышено содержание прямого билирубина (иногда и непрямого), в моче отсутствует стеркобилин, присутствует билирубин, в кале понижено содержание стеркобилина. 26.5.5. Условно физиологическая желтуха новорождённых развивается у большинства здоровых новорождённых в первые дни после рождения и продолжается около двух недель. При различных заболеваниях, возникающих у новорождённых, а также у недоношенных детей желтушный период затягивается. Увеличение длительности гипербилирубинемии может привести к серьёзным последствиям: накоплению билирубина в ткани мозга (ядерная желтуха). Повышению содержания билирубина в крови новорождённых могут способствовать следующие особенности обмена веществ в их организме: · замена фетального гемоглобина на гемоглобин А. В первые дни после рождения усиливается гемолиз эритроцитов, содержващих HbF; образуются новые эритроциты, содержащие HbA. HbF подвергается катаболизму; образуется значительное количество билирубина; · отвлечение альбуминов плазмы для транспорта жирных кислот. Содержание углеводов в организме новорождённых сравнительно невелико; основным энергетическим субстратом являются жирные кислоты, концентрация которых в крови повышается, жирные кислоты транспортируются в комплексе с альбуминами; · низкая активность глюкуронилтрансферазы в ткани печени. Замедление процессов конъюгации билирубина в печени затрудняет его выведение в кишечник; · стерильность кишечника. В кишечнике новорождённого отсутствует микрофлора, поэтому билирубин не превращается в стеркобилиноген и может происходить его обратное всасывание в кровоток. |
| Раздел 26.6 | Нуклеопротеины и их катаболизм в желудочно-кишечном тракте. |
26.6.1. Нуклеопротеины – сложные белки, содержащие в качестве простетической группы нуклеиновые кислоты (РНК или ДНК). В зависимости от того, какая кислота входит в их состав, различают рибонуклеопротеины и дезоксирибонуклеопротеины.
Структурными единицами РНК и ДНК являются нуклеотиды, каждый из которых в свою очередь состоит из азотистого основания и пентозы (образующих нуклеозид), соединённых с остатком фосфорной кислоты.
Пентоза и азотистое основание в каждом нуклеотиде связаны N-гликозидной связью, фосфат и пентоза – сложноэфирной связью. Отдельные нуклеотиды в ДНК или РНК связаны 3’,5’-фосфодиэфирной связью (см. рисунок 26.4).
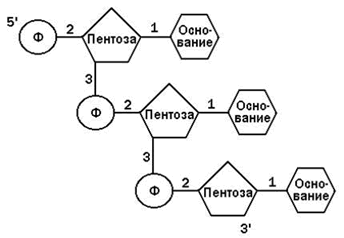 Рисунок 26.4. Схема строения фрагмента полинуклеотидной цепи. Примечание. Пентоза – рибоза в РНК, дезоксирибоза в ДНК. Азотистое основание – аденин, гуанин, цитозин, урацил в РНК, аденин, гуанин, цитозин, тимин в ДНК. Цифрами обозначены типы связей: 1 – N-гликозидные; 2 – сложноэфирные в нуклеотиде; 3 – 3’,5’-фосфодиэфирные (между нуклеотидами).
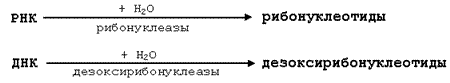
26.6.2. Катаболизм нуклеопротеинов в желудочно-кишечном тракте. Нуклеопротеины, поступающие в организм с пищей, подвергаются расщеплению под действием протеолитических ферментов кишечника. Высвобождаемые нуклеиновые кислоты гидролизуются при участии рибонуклеаз и дезоксирибонуклеаз панкреатического сока до мононуклеотидов. Действие панкреатических нуклеаз дополняют фосфоэстеразы кишечного сока. Схематично это можно представить следующим образом:
Рисунок 26.4. Схема строения фрагмента полинуклеотидной цепи. Примечание. Пентоза – рибоза в РНК, дезоксирибоза в ДНК. Азотистое основание – аденин, гуанин, цитозин, урацил в РНК, аденин, гуанин, цитозин, тимин в ДНК. Цифрами обозначены типы связей: 1 – N-гликозидные; 2 – сложноэфирные в нуклеотиде; 3 – 3’,5’-фосфодиэфирные (между нуклеотидами).
26.6.2. Катаболизм нуклеопротеинов в желудочно-кишечном тракте. Нуклеопротеины, поступающие в организм с пищей, подвергаются расщеплению под действием протеолитических ферментов кишечника. Высвобождаемые нуклеиновые кислоты гидролизуются при участии рибонуклеаз и дезоксирибонуклеаз панкреатического сока до мононуклеотидов. Действие панкреатических нуклеаз дополняют фосфоэстеразы кишечного сока. Схематично это можно представить следующим образом:
 Далее, под воздействием нуклеотидаз и фосфатаз происходит гидролиз нуклеотидов до нуклеозидов, которые либо всасываются, либо под воздействием ферментов слизистой кишечника расщепляются до пуриновых и пиримидиновых оснований.
В организме человека большая часть пуриновых и пиримидиновых оснований, высвободившихся из нуклеиновых кислот, которые поступают с пищей, превращается в конечные продукты (при этом не происходит их включения во вновь образующиеся молекулы нуклеиновых кислот в тканях организма). То есть, нуклеиновые кислоты пищи практически не выступают в роли поставщика непосредственных предшественников нуклеиновых кислот тканей организма.
В то же время нуклеотиды и нуклеозиды, введённые парентерально, могут включаться в нуклеиновые кислоты без всяких изменений. Это послужило основой методов исследования метаболизма нуклеиновых кислот путём введения меченных радиоактивными изотопами азотистых оснований.
Далее, под воздействием нуклеотидаз и фосфатаз происходит гидролиз нуклеотидов до нуклеозидов, которые либо всасываются, либо под воздействием ферментов слизистой кишечника расщепляются до пуриновых и пиримидиновых оснований.
В организме человека большая часть пуриновых и пиримидиновых оснований, высвободившихся из нуклеиновых кислот, которые поступают с пищей, превращается в конечные продукты (при этом не происходит их включения во вновь образующиеся молекулы нуклеиновых кислот в тканях организма). То есть, нуклеиновые кислоты пищи практически не выступают в роли поставщика непосредственных предшественников нуклеиновых кислот тканей организма.
В то же время нуклеотиды и нуклеозиды, введённые парентерально, могут включаться в нуклеиновые кислоты без всяких изменений. Это послужило основой методов исследования метаболизма нуклеиновых кислот путём введения меченных радиоактивными изотопами азотистых оснований.
|
| Раздел 26.7 | Катаболизм нуклеотидов в тканях. |
26.7.1. Нуклеотиды, которые образуются при гидролизе РНК и ДНК, распадаются внутриклеточно.
Нуклеотидазы удаляют фосфат из 5’-положения у пуриновых и пиримидиновых рибо- и дезоксирибонуклеотидов, превращая их таким образом в рибо- и дезоксирибонуклеозиды.
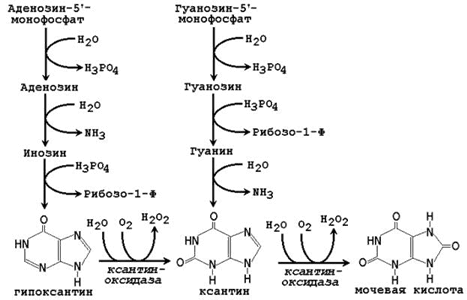 Рисунок 26.5. Катаболизм пуриновых нуклеотидов.
Аденозинмонофосфат (АМФ) подвергается гидролизу с образованием аденозина, последний подвергается дезаминированию и превращается в инозин;
Гуанозинмонофосфат (ГМФ) подвергается гидролизу с образованием гуанозина.
Нуклеозидфосфорилазы катализируют фосфоролиз нуклеозидов с образованием свободных оснований и рибозо-1-фосфата или дезоксирибозо-1-фосфата. При фосфоролизе разрывается N-гликозидная связь при взаимодействии с неорганическим фосфатом.
Конечным продуктом распада пуринов является мочевая кислота, которая образуется следующим путём.
Гипоксантин, продукт распада АМФ, окисляется до ксантина ферментом ксантиноксидазой.
Гуанин, продукт распада ГМФ, дезаминируется с образованием ксантина.
Ксантин окисляется ксантиноксидазой до мочевой кислоты. В реакции окисления с участием ксантиноксидазы используется кислород (О2) и образуется пероксид водорода (Н2О2). Ксантиноксидаза содержит молибден, поэтому этот элемент необходим человеку в небольших количествах. Фермент также содержит железо и серу.
Мочевая кислота и её соли (ураты) плохо растворимы в воде, поэтому в крови они транспортируются частично в комплексе с уратсвязывающим белком. Содержание мочевой кислоты в сыворотке крови здоровых людей составляет 0,24-0,59 ммоль/л у мужчин и 0,16-0,40 ммоль/л у женщин.
Урат натрия легко фильтруется почечными клубочками млекопитающих, интенсивно реабсорбируется в проксимальных канальцах, затем секретируется в петле Генле и, вероятно, снова реабсорбируется в дистальных канальцах. За сутки здоровым человеком выделяется 400 – 600 мг мочевой кислоты. Большое количество фармакологических препаратов и природных соединений оказывает влияние на реабсорбцию урата натрия в почечных канальцах и его экскрецию. Аспирин в больших дозах ингибирует как экскрецию, так и реабсорбцию мочевой кислоты в почках.
26.7.3.Схема катаболизма пиримидиновых нуклеотидов в тканях представлена на рисунке 26.6.
Рисунок 26.5. Катаболизм пуриновых нуклеотидов.
Аденозинмонофосфат (АМФ) подвергается гидролизу с образованием аденозина, последний подвергается дезаминированию и превращается в инозин;
Гуанозинмонофосфат (ГМФ) подвергается гидролизу с образованием гуанозина.
Нуклеозидфосфорилазы катализируют фосфоролиз нуклеозидов с образованием свободных оснований и рибозо-1-фосфата или дезоксирибозо-1-фосфата. При фосфоролизе разрывается N-гликозидная связь при взаимодействии с неорганическим фосфатом.
Конечным продуктом распада пуринов является мочевая кислота, которая образуется следующим путём.
Гипоксантин, продукт распада АМФ, окисляется до ксантина ферментом ксантиноксидазой.
Гуанин, продукт распада ГМФ, дезаминируется с образованием ксантина.
Ксантин окисляется ксантиноксидазой до мочевой кислоты. В реакции окисления с участием ксантиноксидазы используется кислород (О2) и образуется пероксид водорода (Н2О2). Ксантиноксидаза содержит молибден, поэтому этот элемент необходим человеку в небольших количествах. Фермент также содержит железо и серу.
Мочевая кислота и её соли (ураты) плохо растворимы в воде, поэтому в крови они транспортируются частично в комплексе с уратсвязывающим белком. Содержание мочевой кислоты в сыворотке крови здоровых людей составляет 0,24-0,59 ммоль/л у мужчин и 0,16-0,40 ммоль/л у женщин.
Урат натрия легко фильтруется почечными клубочками млекопитающих, интенсивно реабсорбируется в проксимальных канальцах, затем секретируется в петле Генле и, вероятно, снова реабсорбируется в дистальных канальцах. За сутки здоровым человеком выделяется 400 – 600 мг мочевой кислоты. Большое количество фармакологических препаратов и природных соединений оказывает влияние на реабсорбцию урата натрия в почечных канальцах и его экскрецию. Аспирин в больших дозах ингибирует как экскрецию, так и реабсорбцию мочевой кислоты в почках.
26.7.3.Схема катаболизма пиримидиновых нуклеотидов в тканях представлена на рисунке 26.6.
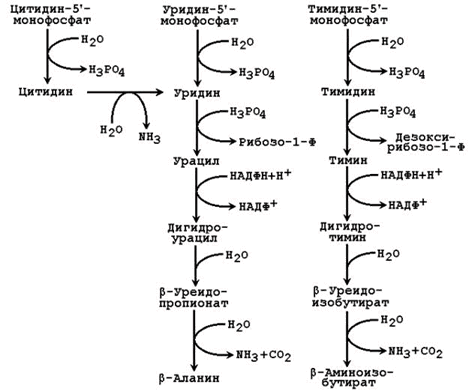 Рисунок 26.6. Катаболизм пиримидиновых нуклеотидов.
Цитидинмонофосфат (ЦМФ) подвергается гидролизу с образованием цитидина, последний вступает в реакцию дезаминирования и переходит в уридин.
Уридинмонофосфат (УМФ) подвергается гидролизу с образованием уридина.
Тимидинмонофосфат (ТМФ) подвергается гидролизу с образованием тимидина.
Нуклеозидфосфорилазы катализируют фосфоролиз нуклеозидов с образованием свободных оснований и рибозо-1-фосфата или дезоксирибозо-1-фосфата.При фосфоролизе разрывается N-гликозидная связь при взаимодействии с неорганическим фосфатом.
Нуклеозидфосфорилазы легко катализируют и обратные превращения свободных оснований в нуклеозиды. Обратные реакции важны для путей реутилизации оснований, особенно урацила и тимина.
26.7.4.Конечными продуктами распада пиримидинов являются β-аланин (образуется из урацила) и β-аминоизобутират (из тимина), которые выводятся с мочой. β-Аланин может служить источником для синтеза коэнзима А, а также ансерина и карнозина – компонентов мышечной ткани.
Рисунок 26.6. Катаболизм пиримидиновых нуклеотидов.
Цитидинмонофосфат (ЦМФ) подвергается гидролизу с образованием цитидина, последний вступает в реакцию дезаминирования и переходит в уридин.
Уридинмонофосфат (УМФ) подвергается гидролизу с образованием уридина.
Тимидинмонофосфат (ТМФ) подвергается гидролизу с образованием тимидина.
Нуклеозидфосфорилазы катализируют фосфоролиз нуклеозидов с образованием свободных оснований и рибозо-1-фосфата или дезоксирибозо-1-фосфата.При фосфоролизе разрывается N-гликозидная связь при взаимодействии с неорганическим фосфатом.
Нуклеозидфосфорилазы легко катализируют и обратные превращения свободных оснований в нуклеозиды. Обратные реакции важны для путей реутилизации оснований, особенно урацила и тимина.
26.7.4.Конечными продуктами распада пиримидинов являются β-аланин (образуется из урацила) и β-аминоизобутират (из тимина), которые выводятся с мочой. β-Аланин может служить источником для синтеза коэнзима А, а также ансерина и карнозина – компонентов мышечной ткани.
|
| Раздел 26.8 | Биосинтез пуриновых нуклеотидов. |
26.8.1.Ключевым соединением в биосинтезе как пуриновых, так и пиримидиновых нуклеотидов является 5-фосфорибозил-1-пирофосфат (ФРПФ). Это соединение участвует также в синтезе коферментов НАД+ и НАДФ+.
ФРПФ образуется при взаимодействии рибозо-5-фосфата и АТФ. Источниками рибозофосфата служат пентозофосфатный путь и распад нуклеотидов. Катализирует реакцию фермент ФРПФ-синтаза.
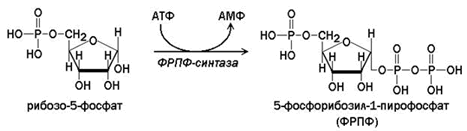 Внутриклеточная концентрация ФРПФ обычно низкая и строго регулируется. Скорость синтеза ФРПФ зависит от наличия субстратов синтеза, особенно рибозо-5-фосфата, и каталитической активности ФРПФ-синтазы, на которую влияют концентрация неорганического фосфата и концентрация АМФ, ГМФ и ИМФ, выступающих в качестве эффекторов.
26.8.2.Молекула ФРПФ служит основой для последующего синтеза пуринового ядра. Источниками атомов углерода и азота являются аминокислоты глутамин, глицин и аспартат, СО2 и два одноуглеродных производных ТГФК – формил-ТГФК и метенил-ТГФК (рисунок 26.7).
Внутриклеточная концентрация ФРПФ обычно низкая и строго регулируется. Скорость синтеза ФРПФ зависит от наличия субстратов синтеза, особенно рибозо-5-фосфата, и каталитической активности ФРПФ-синтазы, на которую влияют концентрация неорганического фосфата и концентрация АМФ, ГМФ и ИМФ, выступающих в качестве эффекторов.
26.8.2.Молекула ФРПФ служит основой для последующего синтеза пуринового ядра. Источниками атомов углерода и азота являются аминокислоты глутамин, глицин и аспартат, СО2 и два одноуглеродных производных ТГФК – формил-ТГФК и метенил-ТГФК (рисунок 26.7).
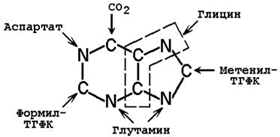 Рисунок 26.7.Происхождение атомов пуринового ядра.
Сначала в реакции, катализируемой фосфорибозил-пирофосфат-амидотрансферазой, из ФРПФ при участии глутамина образуется 5-фосфорибозиламин.
Рисунок 26.7.Происхождение атомов пуринового ядра.
Сначала в реакции, катализируемой фосфорибозил-пирофосфат-амидотрансферазой, из ФРПФ при участии глутамина образуется 5-фосфорибозиламин.
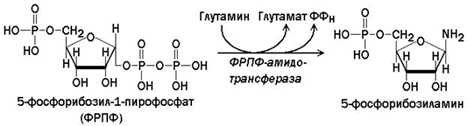 ФРПФ-амидотрансфераза – второй регуляторный фермент синтеза пуриновых нуклеотидов, он ингибируется АМФ и ГМФ по принципу обратной связи. Роль этого фермента в биосинтезе пуринов de novo, однако, менее существенна, чем ФРПФ-синтазы.
Далее к атому азота последовательно присоединяются все остальные компоненты пуринового ядра. Первым продуктом биосинтеза, содержащим готовую пуриновую структуру, является инозинмонофосфат (ИМФ). В его состав входит азотистое основание гипоксантин.
26.8.3. ИМФ является предшественником аденилового и гуанилового нуклеотидов (рисунок 26.5). В синтезе АМФ из ИМФ при взаимодействии аспартатом образуется аденилосукцинат. В следующей реакции отщепляется фумарат и образуется АМФ.
ФРПФ-амидотрансфераза – второй регуляторный фермент синтеза пуриновых нуклеотидов, он ингибируется АМФ и ГМФ по принципу обратной связи. Роль этого фермента в биосинтезе пуринов de novo, однако, менее существенна, чем ФРПФ-синтазы.
Далее к атому азота последовательно присоединяются все остальные компоненты пуринового ядра. Первым продуктом биосинтеза, содержащим готовую пуриновую структуру, является инозинмонофосфат (ИМФ). В его состав входит азотистое основание гипоксантин.
26.8.3. ИМФ является предшественником аденилового и гуанилового нуклеотидов (рисунок 26.5). В синтезе АМФ из ИМФ при взаимодействии аспартатом образуется аденилосукцинат. В следующей реакции отщепляется фумарат и образуется АМФ.
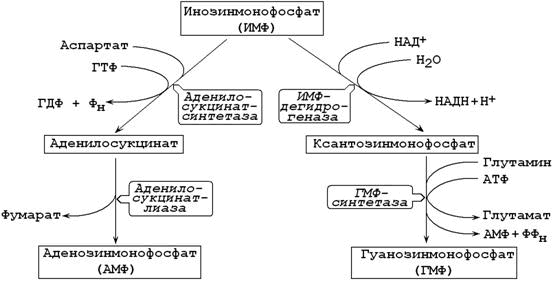 Рисунок 26.8. Образование АМФ и ГМФ из инозинмонофосфата.
Синтез ГМФ из ИМФ также включает две стадии. Сначала ИМФ окисляется в ксантозинмонофосфат, затем добавляется NH2-группа из глутамина.
Интересно отметить, что синтез АМФ требует участия ГТФ, а синтез ГМФ – участия АТФ. Эта особенность биосинтеза способствует поддержанию нужного соотношения адениловых и гуаниловых нуклеотидов в клетке.
26.8.4.Наряду с биосинтезом пуриновых нуклеотидов в клетке de novo существуютпути регенерации пуриновых нуклеотидов из свободных азотистых оснований, образующихся при гидролизе нуклеиновых кислот и нуклеотидов. Эти реакции проще, чем пути синтеза нуклеотидов de novo, и энергетическая цена их значительно меньше. Наибольшее значение имеет механизм фосфорибозилирования пуриновых оснований.
В клетках имеются 2 фермента, участвующих в реакциях синтеза нуклеотидов из пуриновых оснований.
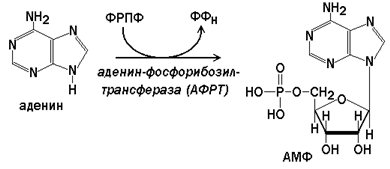
Аденин-фосфорибозилтрансфераза (АФРТ) катализирует перенос фосфорибозы с ФРПФ на аденин:
Рисунок 26.8. Образование АМФ и ГМФ из инозинмонофосфата.
Синтез ГМФ из ИМФ также включает две стадии. Сначала ИМФ окисляется в ксантозинмонофосфат, затем добавляется NH2-группа из глутамина.
Интересно отметить, что синтез АМФ требует участия ГТФ, а синтез ГМФ – участия АТФ. Эта особенность биосинтеза способствует поддержанию нужного соотношения адениловых и гуаниловых нуклеотидов в клетке.
26.8.4.Наряду с биосинтезом пуриновых нуклеотидов в клетке de novo существуютпути регенерации пуриновых нуклеотидов из свободных азотистых оснований, образующихся при гидролизе нуклеиновых кислот и нуклеотидов. Эти реакции проще, чем пути синтеза нуклеотидов de novo, и энергетическая цена их значительно меньше. Наибольшее значение имеет механизм фосфорибозилирования пуриновых оснований.
В клетках имеются 2 фермента, участвующих в реакциях синтеза нуклеотидов из пуриновых оснований.
Аденин-фосфорибозилтрансфераза (АФРТ) катализирует перенос фосфорибозы с ФРПФ на аденин:
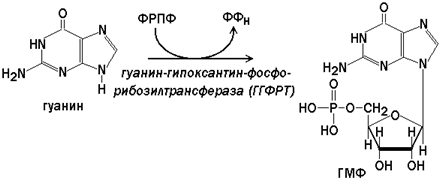
 Гипоксантин-гуанин-фосфорибозилтрансфераза(ГГФРТ) катализирует перенос фосфорибозы с ФРПФ на гуанин или гипоксантин:
Гипоксантин-гуанин-фосфорибозилтрансфераза(ГГФРТ) катализирует перенос фосфорибозы с ФРПФ на гуанин или гипоксантин:
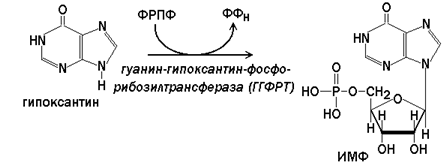 Реакции с участием второго фермента протекают более активно, чем синтез АМФ из аденина.
26.8.5. Нарушения обмена пуриновых нуклеотидов. При нарушениях пуринового обмена часто наблюдается гиперурикемия– повышение содержания мочевой кислоты в крови. Гиперурикемия может быть первичной или вторичной.
Первичная гиперурикемия является ведущим симптомом подагры– полиэтиологического заболевания, как правило, наследственной природы. Гиперурикемия при подагре обусловлена главным образом, избыточным образованием образованием мочевой кислоты, а также снижением её экскреции с мочой. Значительная и длительная гиперурикемия сопровождается отложением солей мочевой кислоты в хрящевой ткани, сухожилиях и слизистых сумках суставов. Накопление кристаллов уратов в тканях может вызывать резкую воспалительную реакцию (подагрический артрит), что приводит впоследствии к деформации сустава. Избыток мочевой кислоты способствует также образованию уратных камней в нижних отделах мочевыводящих путей.
Повышение уровня мочевой кислоты в крови отмечается также при наследственных дефектах некоторых ферментов:
Синдром Леша-Нихана (полное отсутствие ГГФРТ) наследуется как сцепленный с Х-хромосомой рецессивный признак. Болезнь характеризуется параличом, сопровождающимся судорогами, стремлением к членовредительству и тяжёлой гиперурикемией. Вследствие ферментативного дефекта нарушается переход гуанина и гипоксантина в ГМФ и ИМФ соответственно и указанные пуриновые основания превращаются в мочевую кислоту. Кроме того, повышенная концентрация ФРПФ способствует усилению синтеза пуринов de novo. Биохимическая основа неврологических отклонений при синдроме Леша-Нихана неизвестна.
Гликогеноз I типа или болезнь Гирке (дефицит глюкозо-6-фосфатазы) сопровождается повышением активности пентозофосфатного пути и приводит к повышению внутриклеточного уровня рибозо-5-фосфата, из которого синтезируется ФРПФ. Повышенный уровень ФРПФ приводит к увеличению синтеза пуринов de novo. Для данного заболевания характерен также лактатный ацидоз, приводящий к повышению порога секреции уратов почками; это способствует накоплению уратов в организме.
Вторичная гиперурикемия сопутствует заболеваниям, сопровождающимся усиленным распадом клеток (лейкозы, серповидно-клеточная анемия, сахарный диабет, псориаз).
Реже встречается гипоурикемия– снижение содержания мочевой кислоты в крови. Она может быть связана с понижением реабсорбции уратов из клубочкового фильтрата в почках. В этом случае наблюдается увеличение экскреции мочевой кислоты с мочой.
Гипоурикемия развивается и при недостаточности ксантиноксидазы, возникающей при генетическом дефекте фермента или при тяжёлом поражении печени. Это состояние сопровождается повышенной экскрецией гипоксантина и ксантина (ксантинурией), а также образованием в почках ксантиновых камней.
Реакции с участием второго фермента протекают более активно, чем синтез АМФ из аденина.
26.8.5. Нарушения обмена пуриновых нуклеотидов. При нарушениях пуринового обмена часто наблюдается гиперурикемия– повышение содержания мочевой кислоты в крови. Гиперурикемия может быть первичной или вторичной.
Первичная гиперурикемия является ведущим симптомом подагры– полиэтиологического заболевания, как правило, наследственной природы. Гиперурикемия при подагре обусловлена главным образом, избыточным образованием образованием мочевой кислоты, а также снижением её экскреции с мочой. Значительная и длительная гиперурикемия сопровождается отложением солей мочевой кислоты в хрящевой ткани, сухожилиях и слизистых сумках суставов. Накопление кристаллов уратов в тканях может вызывать резкую воспалительную реакцию (подагрический артрит), что приводит впоследствии к деформации сустава. Избыток мочевой кислоты способствует также образованию уратных камней в нижних отделах мочевыводящих путей.
Повышение уровня мочевой кислоты в крови отмечается также при наследственных дефектах некоторых ферментов:
Синдром Леша-Нихана (полное отсутствие ГГФРТ) наследуется как сцепленный с Х-хромосомой рецессивный признак. Болезнь характеризуется параличом, сопровождающимся судорогами, стремлением к членовредительству и тяжёлой гиперурикемией. Вследствие ферментативного дефекта нарушается переход гуанина и гипоксантина в ГМФ и ИМФ соответственно и указанные пуриновые основания превращаются в мочевую кислоту. Кроме того, повышенная концентрация ФРПФ способствует усилению синтеза пуринов de novo. Биохимическая основа неврологических отклонений при синдроме Леша-Нихана неизвестна.
Гликогеноз I типа или болезнь Гирке (дефицит глюкозо-6-фосфатазы) сопровождается повышением активности пентозофосфатного пути и приводит к повышению внутриклеточного уровня рибозо-5-фосфата, из которого синтезируется ФРПФ. Повышенный уровень ФРПФ приводит к увеличению синтеза пуринов de novo. Для данного заболевания характерен также лактатный ацидоз, приводящий к повышению порога секреции уратов почками; это способствует накоплению уратов в организме.
Вторичная гиперурикемия сопутствует заболеваниям, сопровождающимся усиленным распадом клеток (лейкозы, серповидно-клеточная анемия, сахарный диабет, псориаз).
Реже встречается гипоурикемия– снижение содержания мочевой кислоты в крови. Она может быть связана с понижением реабсорбции уратов из клубочкового фильтрата в почках. В этом случае наблюдается увеличение экскреции мочевой кислоты с мочой.
Гипоурикемия развивается и при недостаточности ксантиноксидазы, возникающей при генетическом дефекте фермента или при тяжёлом поражении печени. Это состояние сопровождается повышенной экскрецией гипоксантина и ксантина (ксантинурией), а также образованием в почках ксантиновых камней.
|
| Раздел 26.9 | Биосинтез пиримидиновых нуклеотидов. |
26.9.1.Структура пиримидинового ядра проще, чем структура пуринового, и путь биосинтеза его короче.
Главное отличие от биосинтеза пуринов заключается в том, что сборка пиримидинового ядра не требует участия фосфорибозилпирофосфата (ФРПФ). Пиримидиновая структура сначала образуется, а потом взаимодействует с ФРПФ с образованием нуклеотидов.
Предшественниками атомов углерода и азота пиримидинового кольца являются СО2 и аминокислоты глутамин и аспартат (рисунок 26.9).
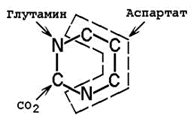 Рисунок 26.9. Происхождение атомов пиримидинового ядра.
26.9.2.При взаимодействии СО2 и глутамина образуется карбамоилфосфат. Реакция протекает в цитозоле с затратой АТФ.
Рисунок 26.9. Происхождение атомов пиримидинового ядра.
26.9.2.При взаимодействии СО2 и глутамина образуется карбамоилфосфат. Реакция протекает в цитозоле с затратой АТФ.
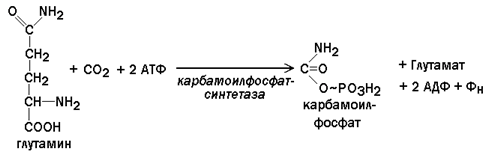 Следует напомнить, что карбамоилфосфат также образуется в процессе синтеза мочевины, но эта реакция происходит только в митохондриях печени и катализируется другим ферментом.
Карбамоилфосфатсинтетаза, участвующая в биосинтезе пиримидинов, - аллостерический фермент, он ингибируется УТФ, пуриновыми нуклеотидами, активируется ФРПФ.
26.9.3.Следующая реакция биосинтеза пиримидинов – образование карбамоиласпартата. Реакцию катализирует аспартат-транскарбамоилаза (АТК-аза).
Следует напомнить, что карбамоилфосфат также образуется в процессе синтеза мочевины, но эта реакция происходит только в митохондриях печени и катализируется другим ферментом.
Карбамоилфосфатсинтетаза, участвующая в биосинтезе пиримидинов, - аллостерический фермент, он ингибируется УТФ, пуриновыми нуклеотидами, активируется ФРПФ.
26.9.3.Следующая реакция биосинтеза пиримидинов – образование карбамоиласпартата. Реакцию катализирует аспартат-транскарбамоилаза (АТК-аза).
 АТК-аза – аллостерический фермент, его ингибитором является цитидинтрифосфат (ЦТФ) – конечный продукт биосинтеза пиримидиновых нуклеотидов.
Дальнейший путь образования пиримидиновых нуклеотидов представлен на рисунке 26.10.
АТК-аза – аллостерический фермент, его ингибитором является цитидинтрифосфат (ЦТФ) – конечный продукт биосинтеза пиримидиновых нуклеотидов.
Дальнейший путь образования пиримидиновых нуклеотидов представлен на рисунке 26.10.
 Рисунок 26.10. Схема синтеза пиримидиновых нуклеотидов.
Рисунок 26.10. Схема синтеза пиримидиновых нуклеотидов.
|
| Раздел 26.10 | Особенности биосинтеза дезоксирибонуклеотидов. |
26.10.1.Дезоксирибонуклеотиды входят в состав ДНК. Содержание дезоксирибонуклеотидов в клетке обычно низкое и повышается только перед репликацией ДНК.
Дезоксирибонуклеотиды образуются в результате восстановления рибонуклеотидов. Эта реакция катализируется рибонуклеотидредуктазой (рисунок 26.11).
В процессе восстановления рибонуклеозиддифосфатов окисляется донор электронов – тиоредоксин. Окисленный тиоредоксин восстанавливается тиоредоксинредуктазой за счёт восстановленного НАДФ.
26.10.2. Тимидинмонофосфат образуется из дезоксиуридинмонофосфата при участии фермента тимидилатсинтазы (рисунок 26.12). Источником метильной группы в реакции служит 5,10-метилен-тетрагидрофолиевая кислота (5,10-метилен-ТГФК), которая окисляется до дигидрофолиевой. Тетрагидрофолиевая кислота регенерируется дигидрофолатредуктазой в реакции, которая требует участия НАДФН. Образование 5,10-метилен-ТГФК происходит в реакции ТГФК с серином или глицином (см. тему 25).
 Рисунок 26.12.Схема образования тимидилового нуклеотида.
Рисунок 26.12.Схема образования тимидилового нуклеотида.
|
| <== предыдущая | | | следующая ==> |
| | |
Date: 2015-07-17; view: 762; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |