Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Молекулярные механизмы и типы генетической рекомбинации
CPЗ 2 часть
1. В зависимости от типа клеток, в которых происходит кроссинговер.
- мейотический - происходит в профазу первого деления мейоза, при образовании половых клеток.
- митотический – при делении соматических клеток, главным образом эмбриональных. Приводит к мозаичности в проявлении признаков.
2. В зависимости от молекулярной гомологии участков хромосом, вступающих в кроссинговер.
- обычный (равный) – происходит обмен разными участками хромосом.
- неравный - наблюдается разрыв в нетождественных участках хромосом.
3. В зависимости от количества образованных хиазм и разрывов хромосом с последующих перекомбинацией генов.
-одинарный
-двойной
-множественный
Значение кроссинговера:
- приводит к увеличению комбинативной изменчивости
- приводит к увеличению мутаций.
2. ПОПУЛЯЦИОННАЯ ГЕНЕТИКА, раздел генетики, изучающий генофонд популяций и его изменение в пространстве и во времени. Разберемся подробнее в этом определении. Особи не живут поодиночке, а образуют более или менее устойчивые группировки, сообща осваивая среду обитания. Такие группировки, если они самовоспроизводятся в поколениях, а не поддерживаются только за счет пришлых особей, называют популяциями. Например, стадо семги, нерестящейся в одной реке, образует популяцию, потому что потомки каждой рыбы из года в год, как правило, возвращаются в ту же реку, на те же нерестилища. У сельскохозяйственных животных популяцией принято считать породу: все особи в ней единого происхождения, т.е. имеют общих предков, содержатся в сходных условиях и поддерживаются единой селекционной и племенной работой. У аборигенных народов популяция – это члены связанных родством стойбищ.
При наличии миграций границы популяций размыты и потому неопределимы. Например, все население Европы – потомки кроманьонцев, заселивших наш континент десятки тысяч лет назад. Изоляция между древними племенами, усиливавшаяся с развитием у каждого из них собственного языка и культуры, вела к различиям между ними. Но обособленность их относительна. Постоянные войны и захваты территории, а в последнее время – гигантская миграция вели и ведут к определенному генетическому сближению народов.
Приведенные примеры показывают, что под словом «популяция» следует понимать группировку особей, связанных территориальной, исторической и репродуктивной общностью.
Закон Харди-Вайнберга
Закон Харди-Вайнберга гласит, что в условиях идеальной популяции частоты генов и генотипов остаются постоянными от поколения к поколению.

Рис. 1
Условия выполнения закона Харди-Вайнберга:
1. Случайность скрещивания в популяции. Это важное условие подразумевает одинаковую вероятность скрещивания между всеми особями, входящими в состав популяции. Нарушения этого условия у человека могут быть связаны с кровнородственными браками. В этом случае в популяции повышается количество гомозигот. На этом обстоятельстве даже основан метод определения частоты кровнородственных браков в популяции, которую вычисляют, определяя величину отклонения от соотношений Харди-Вайнберга.
2. Еще одна причина нарушения закона Харди-Вайнберга - это так называемая ассортативностъ браков, которая связана с неслучайностью выбора брачного партнера. Например, обнаружена определенная корреляция между супругами по коэффициенту интеллекта. Ассортативность может быть положительной или отрицательной и соответственно повышать изменчивость в популяции или понижать ее. Ассортативность влияет не на частоты аллелей, а на частоты гомо- и гетерозигот.
3. Не должно быть мутаций.
4. Не должно быть миграций как в популяцию, так и из нее.
5. Не должно быть естественного отбора.
6. Популяция должна иметь достаточно большие размеры, в противном случае даже при соблюдении остальных условий будут наблюдаться чисто случайные колебания частот генов (так называемый дрейф генов).
Эти положения, конечно, в естественных условиях в той или иной степени нарушаются. Однако в целом их влияние не так сильно выражено, и в человеческих популяциях соотношения закона Харди-Вайнберга, как правило, выполняются.
Закон Харди-Вайнберга позволяет подсчитывать частоты аллелей в популяции. Рецессивные аллели проявляются в фенотипе, если они оказываются в гомозиготном состоянии. Гетерозиготы феноти-пически либо не отличаются от доминантных гомозигот, либо их можно идентифицировать с помощью специальных методов. С помощью закона Ха рди-Вайнберга такой подсчет гетерозигот можно легко сделать по формулам (1) и (2).
Произведем расчеты для рецессивной мутации, вызывающей заболевание фенилкетонурию. Заболевание встречается у одного человека из 10 тысяч. Таким образом, частота встречаемости гомозигот q2 (генотип аа) равна 0,0001. Частота рецессивного аллеля q определяется путем извлечения квадратного корня (q = корень q2) и равна 0,01.
Частота доминантного аллеля будет:
р = 1 -q = 1-0,01 = 0,99.
Отсюда легко определить частоту встречаемости гетерозигот Аа:
2pq = 2 х 0,99 х 0,01 = 0,0198 =0,02, т. е. она составляет приблизительно 2%. Получается, что один человек из 50 является носителем гена фенилкетонурии. Эти данные показывают, какое большое колличество рецессивных генов остается в скрытом состоянии.
3. МОДИФИКАЦИЯ (от лат. modus — мера, вид и facio — делаю)— фенотипическое изменение организма под воздействием факторов внешней среды, Модификации не затрагивают структуру гена, меняется лишь уровень его экспрессии. Чаще всего модификации носят адаптивный характер. Например изменение размера листовой пластинки в зависимости от освещенности. Неадаптивные модификации называются морфозами и представляют собой аномалии или уродства. Предел модификационной изменчивости называется нормой реакции. Чем шире норма реакции, тем выше адаптивные возможности организма. Более широкой нормой реакции обладают количественные признаки, развитие которых определяется несколькими генами (полигенно). Существуют длительные модификации, которые сохраняются на протяжении ряда поколений даже в отсутствии вызвавшего их фактора. Механизм длительных модификаций пока не установлен.
Модификационная изменчивость в теории эволюции
В 1859 году Чарльз Дарвин опубликовал свою работу на эволюционную тему под названием «Происхождение видов путём естественного отбора, или сохранение благоприятных рас в борьбе за жизнь». В ней Дарвин показал постепенное развитие организмов как результат естественного отбора. Естественный отбор состоит из такого механизма:
сначала появляется особь с новыми, совершенно случайными, свойствами (образованными вследствие мутаций)
потом она оказывается или не оказывается способной оставить потомство, в зависимости от этих свойств
наконец, если исход предыдущего этапа оказывается положительным, то она оставляет потомство и её потомки наследуют новоприобретённые свойства
Новые свойства особи формируются вследствие наследственной и модификационной изменчивости. И если наследственная изменчивость характеризуется изменением генотипа и эти изменения наследуются, то при модификационной изменчивости наследуется способность генотипа организмов изменять фенотип при воздействии окружающей среды. При постоянном воздействии одних и тех же условий окружающей среды на генотип могут отбираться мутации, чей эффект сходен с проявлением модификаций, и, таким образом, модификационная изменчивость переходит в наследственную изменчивость (генетическая ассимиляция модификаций). Примером может являться постоянный большой процент пигмента меланина в коже у негроидной и монголоидной расы по сравнению с европеоидной.
Дарвин назвал модификационную изменчивость определённой (групповой).
Определённая изменчивость проявляется у всех нормальных особей вида, подвергшихся определённому воздействию. Определённая изменчивость расширяет пределы существования и размножения организма.
Селекция является одной из важнейших областей практического применения генетики, то есть, генетика – теоретическая основа селекции, так как генетика помогает рационально планировать селекционную работу, исходя из законов наследственности и изменчивости и конкретных особенностей наследования определённого признака.
Кроме этого селекция опирается на достижения других наук, например, систематики и географии растений, цитологии, эмбриологии, биохимии и физиологии растений и животных, молекулярной биологии и др.
Селекция – это наука о методах создания новых и улучшения существующих пород домашних животных и сортов культурных растений и штаммов микроорганизмов.
Селекция – это эволюционный процесс, в котором человек является главным действующим фактором и направляет весь процесс в соответствии со своими потребностями.
Модификация (в селекции) — не связанные с изменением генотипа различия в степени фенотипического проявления одного и того же признака под влиянием меняющихся условий.
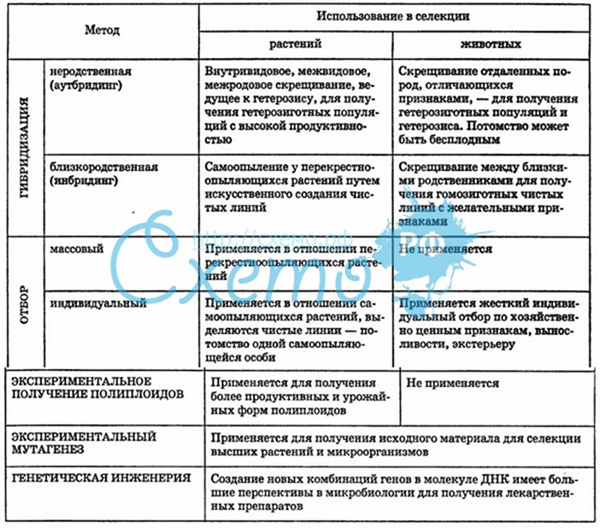
Рис. 2
4. Рекомбинационная изменчивость, генетические рекомбинации.
В основе изменчивости лежит либо изменение реакции генотипа на факторы окружающей среды, либо изменение самого генотипа в результате мутации генов или их рекомбинации. В связи с этим фенотипическую изменчивость подразделяют на наследственную и ненаследственную.
Ненаследственная (средовая, модификационная) изменчивость обусловлена влиянием внутри- и внеклеточных факторов на проявление генотипа. При устранении фактора, вызвавшего модификацию, данные изменения исчезают.
Наследственная (генотипическая) изменчивость, связанная с мутациями, — мутационная изменчивость. Основу мутации составляют изменения последовательности нуклеотидов в ДНК, полная или частичная их утрата, т. е. происходит структурная перестройка генов, проявляющаяся фенотипически в виде измененного признака.
Наследственная изменчивость, связанная с рекомбинациями, называется рекомбинационной изменчивостью.
Молекулярные механизмы и типы генетической рекомбинации
Генетическая рекомбинация - это важный процесс реорганизации генетического материала, обусловленный обменом отдельными сегментами двойных спиралей ДНК, приводящее к возникновению новых комбинаций генов.
Генетическая рекомбинация - главный фактор непостоянства генома, основа большинства его изменений, обусловливающая естественный отбор, микро- и макроэволюции.
Рекомбинация может происходить путем обмена клеточными ядрами, целыми молекулами ДНК или частями молекул. В то время как процессы репликации и репарации ДНК обеспечивают воспроизведение и сохранение генетического материала, рекомбинация приводит к генетической изменчивости.
Она получила развитие у всех живых организмов: у эукариот, у бактерий и даже при размножении вирусов, в том числе таких, генетический материал которых состоит из РНК.
Различают два основных типа генетической рекомбинации:
1) "законную" (общую, или гомологичную), при которой происходит обмен гомологичными (одинаковыми) участками молекул ДНК;
2) "незаконную" (негомологичную), в основе которой лежит обмен негомологичными участками ДНК.
Генетическую рекомбинацию называют сайт-специфичной, если обмен между разными молекулами ДНК осуществляется только в участках со строго определенными нуклеотидными последовательностями,, если в любых местах молекулы ДНК - сайт-неспецифичной.
1. Законная генетическая рекомбинация
Законная генетическая рекомбинация обычно сайт-неспецифична, хотя довольно часто у бактерий и высших организмов она может проявлять черты сайт-специфичности, т. е. избирательности к определенным нуклеотидным последовательностям ДНК (так называемые горячие точки рекомбинации). Такие последовательности резко повышают частоту рекомбинации в тех участках генома, в которых они локализованы.
Гомологичная генетическая рекомбинация
В основе молекулярного механизма законной генетической рекомбинации лежит принцип "разрыв-воссоединение" двух гомологичных молекул ДНК. Этот процесс называют кроссинговер, он включает несколько промежуточных этапов:
1) узнавание участков;
2) разрыв и реципрокное (крест-накрест) воссоединение молекул: замена одних цепей гомологичными;
3) устранение ошибок, возникающих в результате неправильного спаривания участков.
Точка обмена может возникать на любом участке гомологичных нуклеотидных последовательностей хромосом, вовлекаемых в обмен. При этом в точке обмена обычно не происходит изменения нуклеотидных последовательностей. Точность разрыва и воссоединения чрезвычайно велика: ни один нуклеотид не утрачивается, не добавляется и не превращается в какой-нибудь другой.
трансформация (лат. transforma-tio - превращение) - передача ДНК в виде свободного растворимого вещества, выделенного из клетки донора, в клетку реципиента. При этом рекомбинация происходит, если ДНК донора и реципиента родственны друг другу, и может произойти обмен гомологичных участков своей и проникшей извне ДНК. Впервые явление трансформации открыл Ф. Гриффите в 1928 г. Он ввел мышам живой невирулентный бескап-
сульный штамм пневмококка и одновременно убитый вирулентный кап-сульный штамм пневмококка Мыши погибли, из их крови была выделена живая культура вирулентного капсульного пневмококка Сам Гриффите считал, что трансформация произошла путем поглощения невирулентным пневмококком капсульного вещества вирулентного штамма Позже, в 1944 г О Эвери, К Мак Леод и М Мак-Карти доказали, что трансформирующее вещество - это ДНК, которая является носителем генетической информации Гак впервые была доказана роль ДНК как материального субстрата наследственности
трансдукция (лат transductio - перенос) - передача ДНК от бактерии-донора к бактерии-реципиенту с помощью бактериофага Различают неспецифическую трансдукцию, специфическую и абортивную
При неспецифической трансдукции может быть перенесен любой фрагмент ДНК донора При этом ДНК донора попадает в головку бактериофага, не включаясь в его геном Принесенный бактериофагом фрагмент ДНК донора может включиться в хромосому реципиента Таким образом, бактериофаг в этом случае является только переносчиком ДНК, сама фаговая ДНК не участвует в образовании рекомбинанта
При специфической трансдукции гены хромосомы донора замещают собою некторые гены бактериофага В клетке реципиента фаговая ДНК вместе с фрагментом хромосомы донора включается в строго определенные участки хромосомы реципиента в виде профага Реципиент становится лизогенным и приобретает новые свойства
Трансдукция называется абортивной, если фрагмент ДНК, принесенный бактериофагом, не вступает в рекомбинацию с хромосомой реципиента, а остается в цитоплазме и может кодировать синтез какого-то вещества, но не реплцируется при делении, передается только одной из двух дочерних клеток и затем утрачивается.
Роль рекомбинации в эволюции
Эволюционные изменения признаков, детерминируемых одним геном, могут возникнуть в результате сочетания мутационного процесса и отбора. Это сочетание играет наибольшую роль в эволюции бактерий. Оно влияет также на простые признаки многоклеточных организмов, выступая здесь, однако, лишь на вторых ролях в общем процессе изменения.
Эволюция новых сложных признаков у многоклеточных организмов начинается с возникновения изменчивости по множественным генам и завершается закреплением в популяции новой адаптивной комбинации генов. Рекомбинация -- важный промежуточный этап в этом процессе.
Процесс начинается с мутаций в двух или нескольких генах. Мутантные аллели, в случае если они рецессивны, могут оставаться в течение многих поколений в диплоидном состоянии, не экспрессируясь. Диплоидное состояние представляет собой хранилище для мутационной и полигенной изменчивости, а ключом к нему служит половое размножение; оно создаёт всевозможных рекомбинантов из имеющегося в генофонде «сырья»*.
Биологическая функция пола состоит в производстве множества рекомбинантных типов. Вероятность того, что какая-либо определённая комбинация генов может быть собрана в одном ряду поколений за счет одного лишь мутационного процесса и без полового размножения, практически равна нулю. Рассмотрим возникновение производного генотипа abc от предкового генотипа ABC у гаплоидного организма. Если бы это превращение зависело от ряда мутаций в бесполых линиях, то оно протекало бы крайне медленно. Однако в популяции организма с половым размножением новый генотип abc может образоваться всего за два поколения в результате скрещивания между трёмя линиями, каждая из которых несёт по одному аллелю -- a, b или с (Wright, 1931; Miller, 1932*).
Процесс рекомбинации слеп в отношении адаптивной ценности образующихся рекомбинантов. Он механически создаёт как негодные, так и полезные в адаптивном смысле типы рекомбинантов. Совершенно очевидно, что желательно свести долю первых до минимума. В любом сложном организме чужие гены из отдаленно родственной популяции вряд ли могут гармонично сочетаться с коадаптированными генами нативной популяции. Один из путей снижения доли плохо адаптированных рекомбинантов состоит поэтому в возведении преград, препятствующих широкой гибридизации.
5. Под эпигенетикой обычно понимают область знаний о совокупности свойств организма, которые не прямо, а опосредованно закодированы в геноме и, по определению, должны передаваться по наследству. По сути дела в первую очередь эта наука имеет дело с механизмами, контролирующими экспрессию генов и клеточную дифференцировку. У организмов существуют мощные регуляторные элементы (в самом геноме и даже целые системы в клетках), которые контролируют работу генов, в том числе и в зависимости от разных внутренних и внешних сигналов биологической и абиотической природы. Эти сигналы накладываются на генетику и часто по-своему решают коренной вопрос "быть или не быть". Действительно, даже самая отличная генетика может вовсе и не реализоваться, если эпигенетика неблагополучна. По образному выражению Нобелевского лауреата П. Медавара "генетика полагает, а эпигенетика располагает".
Набор и природа эпигенетических сигналов в клетке весьма разнообразны, таких сигналов много и сегодня они разделяются, по крайней мере, на несколько групп - метилирование и деметилирование ДНК, гистоновый код (энзиматическая модификация гистонов - ацетилирование, метилирование, убиквитилирование, фосфорилирование и другие), транскрипционное и трансляционное замалчивание генов малыми РНК, позиционирование элементов хроматина. Любопытно, что многие из этих процессов переплетены между собой и взаимозависимы. Это во многом обеспечивает и гарантирует надежность эпигенетического контроля за избирательным функционированием генов.
Эпигенетическая наследственность - "надгенетическая" передача приобретенных признаков потомству без изменения кода ДНК
Недавно открыто еще несколько способов передачи по наследству приобретенных признаков. Эти способы не связаны с изменениями самого "текста", записанного в структуре молекул ДНК, то есть с мутациями. Поэтому такую наследственность называют "эпигенетической", или "надгенетической".
Один из таких "эпигенетических" механизмов – метилирование ДНК. Оказалось, что в процессе жизнедеятельности к молекулам ДНК в клетках (в том числе и в половых) специальные ферменты "пришивают" метильные группы (-CH3). Причем к одним генам метильных групп пришивают больше, к другим – меньше. Распределение метильных групп по генам ("паттерн метилирования") зависит от того, насколько активно тот или иной ген используется. Получается совсем как с "упражнением" и "неупражнением" органов, которое Ламарк считал причиной наследственных изменений. Поскольку "паттерн метилирования" передается по наследству, и поскольку он, в свою очередь, влияет на активность генов у потомства, легко заметить, что здесь может работать совершенно ламарковский механизм наследования: "натренированные" предками гены будут и у потомства работать активнее, чем "ослабевшие" от долгого неиспользования.
Другой вариант "эпигенетического" наследования приобретенных признаков основан на системах взаимной активации и инактивации генов. Допустим, ген А производит белок, одно из действий которого состоит в блокировании работы гена Б, а ген Б, в свою очередь, кодирует другой белок, способный "выключать" ген А. Такая система может находиться в одном из двух состояний: либо ген А работает, и тогда ген Б выключен, либо наоборот. Допустим, что переход системы из одного состояния в другое может происходить только в результате какого-то особенного внешнего воздействия, и случается такое редко. То состояние, в котором находится эта двухгенная система в клетках матери, будет через яйцеклетку передаваться ее потомству (поскольку сперматозоид содержит пренебрежимо малое количество белков). Если же в течение жизни матери система переключится в другое состояние, то этот приобретенный признак передастся потомству, родившемуся после "переключения".
6.Наследственные болезни — заболевания человека, обусловленные хромосомными и генными мутациями. Нередко ошибочно термины «наследственная болезнь» и «врожденная болезнь» употребляются как синонимы, однако врожденными болезнями называют те заболевания, которые имеются уже при рождении ребенка и могут быть обусловлены как наследственными, так и экзогенными факторами. Таковы, например, пороки развития, связанные с воздействием на эмбрион и плод ионизирующего излучения, химических соединений, лекарственных средств, принимаемых матерью, а также внутриутробных инфекций.
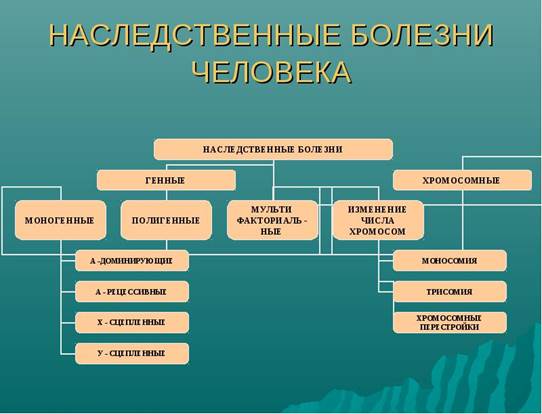
Рис. 3
Идеально отлаженную работу всего организма легко можно увидеть простым глазом: для этого только нужно посмотреть на человека. О том, что у него всё в порядке, скажет цветущий, жизнерадостный и здоровый вид. Дискомфортное и болезненное состояние также легко распознать. В этом случае для несчастного и свет не мил, а прелести жизни отходят на второй план.
Наследственные заболевания
Так живёт нормальный человеческий организм: здоровье чередуются с болезненным состоянием. Его помогают вылечить врачи, и всё опять нормализуется. Но есть люди, к которым болезни прицепляются с самого рождения. Медицина ничем не может помочь таким несчастным, и страшный крест приходится нести до конца жизни.
Разговор идёт о наследственных заболеваниях, причина которых кроется в генетическом аппарате человека. Он очень надёжен, но как и любой механизм не застрахован от сбоев и поломок. Составляющие этого аппарата – гены, молекулы ДНК, хромосомы. У некоторых людей они могут мутировать, а заложенные в них программы сбиваться.
У взрослого человека «плохие» гены могут никак себя не проявлять. Организм такого индивидуума кажется абсолютно здоровым, без каких либо отклонений. Все аномалии скрыты от посторонних глаз, а носители разрушительной информации ведут себя очень тихо и молчат. Но это обманчивая тишина – затишье перед бурей. Начинается же буря тогда, когда происходит оплодотворение яйцеклетки.
Как каждый знает ещё со школьной скамьи – вся наследственная программа будущего человека распределена в равной пропорции между половой клеткой отца и матери. У женщина это яйцеклетка, у мужчины сперматозоид. Последний, при оплодотворении, проникает в яйцеклетку, растворяя её оболочку специальным ферментом. Ядра половых клеток сливаются и образуется новая, оплодотворённое клетка.
Она начинает делиться. Из одной клетки получается две, потом четыре, восемь и так далее. Зародыш растёт, поудобнее устраивается в теле матери. Через девять месяцев на свет появляется новый человек, который несёт в себе наследственные признаки обоих родителей.
На генетическом уровне эти признаки концентрируются в хромосомах, каждая из которых является универсальной единицей наследственной информации. Хромосом в оплодотворённой клетке 46. Одна половина из них раньше находилась в яйцеклетке, вторая в сперматозоиде. Собравшись в одном ядре, они разбиваются на 23 пары.
Наследственные заболевания человека обусловлены мутациями. Выделяют три основные группы наследственных заболеваний:
1) Хромосомные болезни
2) Генные болезни
3) Мультифакторные заболевания
Хромосомные болезни – наследственные заболевания, которые обусловлены геномными (изменение числа хромосом) и хромосомными (изменение структуры хромосом) мутациями. Хромосомные болезни, как правило, не передаются потомству и встречаются в семьях как спорадичные случаи.
Основная причина возникновения хромосомных болезней – нерасхождение хромосом в мейозе во время гаметогенеза у одного из родителей. Они возникают вследствие мутаций в гаметах одного из здоровых родителей или в зиготе на первых стадиях дробления. Если мутация, возникшая в гаметах, – это полная форма болезни, то на стадии дробления зиготы – мозаичная форма болезни. В отличие от генных, хромосомные мутации охватывают значительно больший объем генетического материала и характеризуются множественными поражениями. Именно они вызывают около 45 % случаев гибели плода после имплантации и 60-70 % – 2-4-недельных выкидышей. Больные хромосомными болезнями занимают почти 25 % госпитализированных пациентов в мире.
Для диагностики хромосомных болезней применяют цитогенетический метод генетики. Количественные и структурные нарушения хромосом определяют под микроскопом.
Хромосомные аномалии встречаются в 1 % новорожденных. Они являются причиной 45-50 % множественных врожденных пороков развития, около 36 % случаев глубокой умственной отсталости, 50 % аменорей у женщин, 10 % бесплодия у мужчин.
Различают хромосомные болезни, обусловленные:
1) изменением количества и структуры аутосом;
2) изменением количества половых хромосом.
1. Хромосомные заболевания, обусловленные изменением количества и структуры аутосом
Синдром Дауна (трисомия 21).
Клиническую картину синдрома впервые в 1866 г. описал английский врач Л. Даун. В 1959 г. французский ученый И.Лежен обнаружил в кариотипе больных лишнюю хромосому 21. Кариотипы больных – 47, ХХ, 21+ или 47, ХУ, 21+. Частота 1:1100, а в некоторых регионах – 1:700-1:800 новорожденных. Риск рождения детей с синдромом Дауна возрастает с возрастом матери. На частоту их рождения не влияют половые, расовые, географические и популяционные отличия. Комплекс врожденных пороков развития, характерных для синдрома Дауна, обуславливает клиническую картину “все дети из одной семьи”.
Клинические диагностические признаки: низкий рост, разная степень умственной отсталости, черепно-лицевые аномалии: косой разрез глаз, короткая шея, эпикант, плоское лицо, маленький короткий нос, большой язык, маленькие деформированные уши. Характерные также мышечная гипотония, разшатанность суставов, поперечная складка на ладонях, клинодактилия мизинца. Врожденные недостатки внутренних органов (сердца), сниженный иммунитет часто являются причиной смерти этим детей.
Цитогенетические варианты синдрома разнообразные. Основную часть (94 %) составляют случаи полной трисомии 21 как результат нерасхождения хромосом в мейозе. При этом взнос материнского нерасхождения составляет 80%, отцовского – 20%. Приблизительно 4% больных имеют транслокационную форму (транслокация хромосомы 21 чаще всего на хромосомы 13 или 22) и 2 % – мозаицизм вследствие митотичного нерасхождения, когда одна часть клеток имеет нормальное количество хромосом (46), а другая – анэуплоидную (47). Транслокационная форма не зависит от возраста матери, поэтому есть высокий риск повторного рождения больного ребенка в семье.
Синдром Патау (трисомия-13).
Кариотип 47, ХХ, 13+ или 47, ХУ, 13+. Частота 1:5000 - 1:7000 новорожденных. Цитогенетические варианты разные. Полная трисомия-13 как следствие нерасхождения хромосом в мейозе в одного из родителей встречается в 80-85 % больных. Другие случаи обусловлены робертсоновскими транслокациями типа D/13 и G/13. Мозаицизм встречается редко.
Клинические диагностические признаки: щели верхней губы и неба («заячья губа» и «волчья пасть»), уменьшенный объем черепа, перекошенный, низкий лоб, микрофтальмия, анофтальмия (отсутствие одного или обеих глазных яблок), переносица запавшая, деформированные ушные раковины, полидактилия; врожденные пороки сердца, других внутренних органов. Большинство детей умирает в первые недели или месяцы. Решающим в диагностике является цитогенетическое исследования.
Мультифакториальные заболевания
Мультифакториальные болезни, или болезни с наследственной склонностью, обусловленные комбинацией генетических и негенетических факторов (внешняя среда).
Для реализации мультифакториальных болезней необходима не только соответствующая генетическая конституция индивида, но и фактор или комплекс факторов среды, которые сыграют роль пусковых моментов в формировании патологии. К таким заболеваниям относятся: атеросклероз, подагра, ревматизм, ишемическая болезнь сердца, гипертоническая болезнь, эпилепсия, язвенная болезнь желудка и 12-палой кишки, цирроз печени, сахарный диабет, бронхиальная астма, туберкулез, псориаз, шизофрения. Характерные признаки мультифакториальных болезней:
1) большой полиморфизм клинических форм и индивидуальных проявлений; существование переходных форм от здоровых людей к больным, от субклинических форм к тяжелому протеканию болезни;
2) высокая частота в популяции (сахарным диабетом страдает 5 % людей земного шара, аллергическими заболеваниями – свыше 10 %, шизофренией – 1 %, гипертонией – около 30 %);
3) несоответствие наследования законам Менделя;
4) разный возраст больных.
По наследственности передается склонность к определенному заболеванию. Для некоторых клинических форм роль наследственного (семейного) фактора является решающей. Степень риска для родственников больного зависит от частоты болезни в популяции. Чем ближе степень родственности с больным у родственников, тем большая вероятность рождения у них больного ребенка. В ряде случаев наблюдается неодинаковая частота патологии в зависимости от пола. Например, врожденная дисплазия тазобедренного сустава чаще встречается у девочек, а пилоростеноз – у мальчиков.
Болезни с наследственной предрасположенностью могут быть моногенными и полигенными. Основу составляет полигенное наследование и часто гетерозиготность. При полигенном наследовании признак обуславливают несколько неаллельных генов, но проявляются они в зависимости от условий среды. При гетерозиготном носительстве патологический рецессивный ген в гетерозиготном состоянии не проявляется, но может проявиться при неблагоприятных условиях жизни. Поскольку болезни с наследственной склонностью определяются объединением наследственных и внешних факторов, их относят к заболеваниям с пенетрантностью, которая в значительной мере зависит от условий среды. Изменяя условия среды, можно значительно изменить проявление таких болезней и даже предупредить их.
Генные болезни - разнородная по клиническим проявлениям группа заболеваний, обусловленных мутациями на генном уровне. Основой для объединения их в одну группу служат этиологическая генетическая характеристика и соответственно закономерности наследования в семьях и популяциях. Коль скоро мутации в индивидуальных генах являются этиологическим фактором генных болезней, то закономерности их наследования соответствуют менделевским правилам расщепления в потомстве, т.е. формальная генетика генных наследственных болезней ничем не отличается от «поведения» в семьях любых менделирующих признаков. «Поведение» некоторых патологических генов может отклоняться от менделевско-моргановских правил в связи с фенотипическими эффектами (летальность, стерильность). Необходимо, однако, сразу сделать пояснения в отношении содержания понятий «генные мутации» и «менделирующая наследственность» у человека.
Во-первых, согласно многочисленным исследованиям разных наследственных болезней и генома человека в целом, можно говорить о многообразии видов мутаций в одном и том же гене, которое является причиной наследственных болезней. У человека описаны виды генных мутаций, обусловливающие наследственные болезни: миссенс, нонсенс, сдвиг рамки считывания, делеции, вставки (инсерции), нарушения сплайсинга, увеличение числа (экспансия) тринуклеотидных повторов. Любой из этих видов мутаций может вести к наследственным болезням. Даже одна и та же генная болезнь может быть обусловлена разными мутациями. Например, в гене муковисцидоза описано около 200 вызывающих болезнь мутаций (всего их около 1000) следующих типов: делеции, миссенс, нонсенс, сдвиг рамки считывания, нарушения сплайсинга. Для гена фенилкетонурии известно более 30 патологических мутаций (миссенс, нонсенс, делеции, нарушения сплайсинга).
7. Мобильные генетические элементы (МГЭ, подвижные элементы, транспозируемые элементы, транспозоны и т.д.) повсеместно распространены в живой природе от плазмид, фагов и бактерий до высших животных и растений (Хесин, 19S4; Berg, Howe, 1989). Будучи нестабильными по своей локализации в геномах, они создают мощный источник изменчивости генов, систем их управления и геномов (Lambert et al., 1988). Будучи сами последовательностями нуклеотидов, они тоже подвержены эволюции. Поэтому МГЭ выступают и как факторы эволюции содержащих их геномов, и как эволюционирущие объекты.[3,c.4]
I. МГЭ как факторы изменчивости
В настоящее время МГЭ найдены у бактерий (включая их фагов и плазмид), низших грибов, насекомых, растений, животных и многих других объектов (Хесин, 1984; Berg, Howe,1989). Число известных семейств МГЭ, вероятно превышает 100. У хорошо изученных объектов найдены многие десятки семейств МГЭ: например у дрозофилы их число, вероятно, достигает 50 (Berg, Howe, 1989; Charlesworth, Langley, 1989). Некоторые семейства относятся к умеренным повторам, имея десятки-сотни копий на геном, другие - к повторам высокой множественности (Alu у человека - 5--6 млн копий на геном (Fa-Ten-Kao, I985)). В сумме МГЭ различных семейств могут составлять значительную часть генома (до 10% у дрозофилы (Finnegan, 1985; Ананьев, 1989)). МГЭ нестабильны, т.е. с определенной вероятностью способны к транспозициям и исключению из отдельных позиций генома (табл.1).
МГЭ способны к воспроизведению в клетке либо через репликацию ДНК, либо через прямую и обратную транскрипцию (ретротранспозоны).
Date: 2016-05-25; view: 2598; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |