Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Жасушааралық сигнализация және интеграцияның молекулярлық механизмдері
- Жасушаға сыртқы биомолекулярлық сигналдарды беру жолдары және жасушаның жауап беру механизмдері.
Жасуша жиі өзіне тән ерекше сигнал жауабына молекулярлы каскадты механизмді жібереді, ол биохимиялық жауап кезінде интегративті жол жүзеге асырылады
• Жасушалық жауаптың ерекшелігі рецептор түрімен анықталынады. Рецептор- нақты заттарға (лигандаларға) сәйкестігі бар, ақуыз молекуласы. Лигандалар (нейромедиаторлар,гормондар, өсу факторлары, дәрі – дәрмектер, т.б.)
• 1- жасушада (беткейлік, немесе трансмембраналық),
• 2- цитозолде (цитоплазмалық),
• 3- ядрода.
• Олар біріншіден ерекше сигналдарды алу үшін, екіншіден жасушалық
жауапты шақыратын каскадтық ферментативтік реакцияларды жіберу үшін
қызмет атқарады.
• Сигналды берудің жалпы кестесінің 5 негізгі этаптары бар.
•. 1 этап – Интегральдық ақуыздың немесе рецептордың лигандамен байланысуы (оларға гормондар,дәрі-дәрмек, немесе антидене жатады).
• 2 этап – Интегральдық ақуыздың- рецептордың белсенділігі, яғни конформациялық өзгеріске ұшырауы.
• 3 этап – Эктодомен сигналды түрлендіреді. Интегральдық ақуыздың- рецептордың конформациялық өзгерісінен кейін, цитоплазмалық бөлігі эффекторды белсендендіреді.
• 4 этап - эффектор активациясы бұл G-ақуыздың -альфа суббірлігі.
• 5 этап – сигналдың әлсіреуі. Немесе эффектордың қайта белсенсізденуі.
Сигналды берудің бұзылуы әрбір этапта болуы мүмкін, ол ауруларға және патологиялық процесстерге әкеледі:
- Жасушаның беткейлік рецепторларымен байланысты өтетін сигналдардың берілу механизмдері
- Беткейлік рецепторлармен байланыспай берілетін сигналдардың механизмі
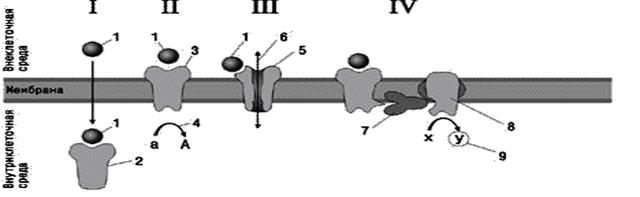
I – сигналдың беткейлік рецепторлармен байланысты емес берілуі, яғни цитоплазмалық рецепторлар арқылы берілуі.
II – сигналдың трансмембраналық рецепторлар арқылы берілуі.
III – сигналдың иондық каналдардың ашылуы немесе жабылуы арқылы берілуі.
IV – сигналды екінші ретті мессенджерлер арқылы берілуі.

G-ақуыздарды ашқан және зерттегендер Альфред Гилман және Мартин Родбелл, оларға осы жаңалықтары үшін физиология және медицина бойынша Нобель силығы 1994 жылы берілген.

• 



• Нысана ретінде, a-суббірлік – ГТФ комплексі үшін, мембранамен байланысқан ферменттер (аденилатциклаза, фосфолипаза С) және иондық каналдар бола алады.
•  2. Жасушалық сигнализация.
2. Жасушалық сигнализация.
• Фосфорлену және дефосфорлену – жасушаішілік сигналды берудегі негізгі механиздері. Бұл процесстерге екі түрлі ферменттер қатысады – ол киназалар және фосфатазалар. Олар сигналдық процесстерде маңызды роль атқарады.
• Фосфорлену және дефосфорлену –бұл қайтымды пайда болатын және фосфатты эфирлі байланыстардың гидролизі.
Биологиялық мембрананың құрылуына трансляцияда, генетикалық материалдың пайда болуында маңызы зор.
• Фосфорленудің мәні
• Фосфорлену – сигналды беру процессінде негізгі 2 - қызметті атқарады:
1- ақуыздардың конформациясының өзгеруі және ферменттердің активациясы.
2- ақуыз молекулаларындағы қосылысу бөліктерін жасайды, яғни белсенденген элементтермен бірлесетін жаңа ақуыздарды тартады, уақытша жасушаішілік берушілердің пайда болуына әкеледі.
• Дефосфорленудің мәні
Кейде фосфатты алып тастау сигналды өшіргеннен кейін немесе сигналды
жолдардың активациясынан кейін жүреді.
• Екіншілік (қосымша) мессенджерлер (тасымалдаушылар ) – бұл кішкентай молекулалар, рецептор активациясына жауап беруде, толып жатқан көп және жылдам жасушада синтезделінеді, және молекулярлық сигналдың күшеюі үшін қызмет атқарады.
•
• Бес қосымша мессенджерлер(аралық көмекші тасмалдаушы) бар, жасушаішілік сигнализация процесінде үлкен маңызды роль атқарады.
• 1 – циклдік аденозинмонофосфат (сАМР)
• 2 – циклдік гуанозинмонофосфат (сGМР)
• 3 – диацилглицеролтрифосфат (DАG)
• 4 – инозитолтрифосфат (ИФ)
• 5 – кальций (Са)
• Бірінші жағдай. Аденилатциклазаны белсендендіруге әкелетін механизм.
• 1. Гормонның рецептормен байланысуы рецептордың конформациясының өзгеруіне және G-белокқа байланысушылығының жоғарылауына әкеледі. G-белоктың 3 протомері (a, b, g-G-белок) және ГДФ комплекс түзеді. Нәтижесінде, гормон-рецептор-G-белок-ГДФ-тен тұратын үштік комплекс пайда болады.
• 2. G-ақуызындағы a-протомердегі ГДФ ГТФ-қа ауыстырылады. Бұл комплекс a --суббірлік-ГТФ және бетта-гамма - димерге диссоциацияланады.
• 3. a --суббірлік-ГТФ мембранамен байланысқан фермент аденилатциклазамен байланысады. Бұл аденилатциклазаның конформациясын өзгертіп, ферментті белсендендіреді. Аденилатциклазаның субстраты АТФ, ал реакциясының өнімі циклдік АМФ болып табылады. Нәтижесінде АТФ-тан циклдік-АМФ-тың түзілу жылдамдығы жоғарылайды.
• 4. циклдік-АМФ молекулалары жасуша ішіндегі көптеген ақуыздармен қайтымды байланыса алады, нәтижесінде ондағы биохимиялық процестердің жылдамдықтары мен бағыттары өзгереді.
• цАМФ ядроға өтіп CREB (cAMP response element binding protein) ақуыздарды белсендіреді. Бұл ақуыздар кейбір гендердің реттеуші элементтерімен арнайы байланыса алады және транскрипция процессін босата алады.
5. a - суббірлік – ГТФ-тің аденилатциклазамен байланысуы, өз кезегінде, a -суббірліктің конформациясын өзгертеді. ГТФ ГДФ-ке ауыстырылады.
• 6. a - суббірлік - ГДФ gb - димермен бірігеді. Нәтижесінде, G-белоктардың 3 протомерінің жиналуы өтеді.
• Екінші жағдай. Бұл жағдайда молекулалық нысана қызметін фосфолипаза С атқарады (инозитолфосфаттық жүйе).
• Фосфолипаза С-ны (инозитолфосфаттық жүйені) белсендендіруге әкелетін механизм.
• 1. Гормонның рецептормен байланысуы оның конформациясын өзгертіп G ақуызға байланысушылығын жоғарлатады.
• 2. Бұл a - суббірліктегі ГДФ-тің ГТФ-ке ауыстырылуына әкеледі.
• 3. G-белок a - суббірлік + ГТФ және gb - димерге ыдырайды
• 4. a - суббірлік + ГТФ мембраналық фермент фосфолипаза С-мен әсерлесіп оны активтендіреді.
• Бұл ферменттің субстарты фосфатидилинозитолбисфосфат (ФИФ) болып табылады. Реакция өнімі: инозитол – 3 фосфат (цитозолға бөлініп шығады) және диацилглицерин (ДАГ) (мембранада қалатын) болып табылады.
• 5. Инозитол-3-фосфат эндоплазмалық ретикулумның спецификалық Са-каналдық орталықтарымен байланысады. Нәтижесінде, оның конформациясы өзгеріп, ол ашылады. Са иондары цитозолға өтеді. Цитозолда инозитол-3-фосфат болмаған кезде, канал жабық болады.
• 6. Цитоплазмадағы С-иондары концентрациясының жоғарылауы, оның
1. белсенсіз цитозолдық фермент протеинкиназа С -мен;
2. кальмодулин ақуызымен әсерлесуіне әкеледі.
• 7. Плазмалық мембранада фермент протеин киназа С (ПКС) болады. Оның белсенденуі үшін оған диацилглицериннің, фосфатидилсериннің (ФС) және Са2+ бірігуі қажет.Фермент белсенденгенген соң жасушадағы әртүрлі ақуыздардың активтіліктері өзгереді.
• 8. Кальмодулин ақуызында Са-иондарын байланыстыратын 4 орталығы болады. Комплекс кальмодулин + 4 Са-иондары + өзі ферменттік белсенділікті көрсетпейді, бірақ цитозолдағы әртүрлі ақуыздар мен ферменттерді белсендендіреді.
• 9. Жасушадағы Са2+ концентрациясын бастапқы деңгейге дейін төмендету үшін Са2+-АТФаза және транслоказа (антипорт) жүйелері қызмет атқарады.
• 10. Инозитол-3-фосфат және ДАГ бірнеше реакциялардан соң мембранадағы фосфатидилинозитолбисфосфат (ФИФ-ке қайта айналады). Белсенді протеин киназа С (ПКС) ФИФ түзілуін күшейтеді.
Date: 2016-05-14; view: 3637; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |