Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Регуляция транскрипции. Теория Оперона
Оперон — участок ДНК, кодирующий строение одного вида белков, содержащий регуляторную зону, контролирующую синтез этих белков.
Регуляция транскрипции м-РНК включает индукцию и репрессию генов.
Оперон состоит из гена-регулятора, гена-промотора, гена-оператора, структурных генов.
Индукция — «включение» процесса транскрипции. Трансляция возможна, если ген-оператор не связан с белком-регулятором. В этом случае РНК-полимераза присоединяется к гену-промотору и начинает синтез РНК, комплементарной структурным генам (м-РНК).
Репрессия — «выключение» транскрипции, когда ген-оператор связан с белком-регулятором и РНК-полимераза не имеет возможности присоединиться к гену-промотору.
Сродство белка-регулятора к гену-оператору может меняться при взаимодействии его с эффекторами.
Индукторами транскрипции служат субстраты метаболических путей, репрессорами, как правило, являются конечные продукты метаболических путей.
Роль индукторов и репрессоров могут играть гормоны.
В норме содержание общих НК в сыворотке крови взрослого человека составляет 80—100 мкг/мл. Увеличение концентрации НК в сыворотке крови свидетельствует об усилении процессов катаболизма в клетках органов и тканей.
Катаболизм белков 3 этапа:
1. Переваривание и всасывание (1% энергии).
2. Межуточный обмен (специфические пути катаболизма) (29% энергии).
3. Общий конечный путь распада — терминальное окисление (70% энергии).
Специфические пути включают в себя гликолиз, b-окисление ВЖК, а также дезаминирование, переаминирование и декарбоксилирование аминокислот.
Общий конечный путь распада включает окислительное декарбоксилирование ПВК, цикл Кребса и ЦПЭ.
Окислительное декарбоксилирование ПВК протекает аэробно в матриксе митохондрий под влиянием мультиэнзимного комплекса (3 фермента и 5 коферментов, М = 10 млн.) в 4 стадии.
Пируватдегидрогеназный комплекс присоединен к внутренней мембране митохондрий со стороны матрикса. Суммарная реакция:
CH3

 | пируватдегидрогеназныйO
| пируватдегидрогеназныйO
 C=O комплекс CH3 – C ~ SKoA+ CO2
C=O комплекс CH3 – C ~ SKoA+ CO2
|
СОООН
Переваривание жира начинается в желудке, где находится малоактивный фермент желудочная липаза, однако ее роль в гидролизе пищевых триглицеридов у взрослых людей невелика. Во-первых, в желудочном соке взрослого человека и других млекопитающих содержание липазы крайне низкое. Во-вторых, рН желудочного сока далек от оптимума действия этого фермента (оптимальное значение рН для желудочной липазы 5,5-7,5). В-третьих, в желудке отсутствуют условия для эмульгирования триглицеридов, а липаза может активно действовать только на триглицериды, находящиеся в форме эмульсии. Поэтому у взрослых людей не эмульгированные триглицериды составляющие основную массу пищевого жира, проходят через желудок без особых изменений.
Расщепление триглицеридов в желудке взрослого человека невелико, но оно в определенной степени облегчает последующее переваривание в кишечнике. Даже незначительное по объему расщепление триглицеридов в желудке приводит к появлению свободных жирных кислот, которые подвергаясь всасыванию в желудке, поступают в кишечник и способствуют там эмульгированию жиров, облегчая таким образом воздействие на них липазы панкреатического сока.
После того как химус попадает в двенадцатиперстную кишку, прежде всего происходит нейтрализация попавшей в кишечник с пищей соляной кислоты желудочного сока бикарбонатами, содержащимися в панкреатическом и кишечном соках. Выделяющиеся при разложении бикарбонатов пузырьки углекислого газа способствуют хорошему перемешиванию пищевой кашицы с пищеварительными соками. Одновременно начинается эмульгирование жира. Наиболее мощное эмульгирующее действие на жиры оказывают соли желчных кислот, попадающие в двенадцатиперстную кишку с желчью в виде натриевых солей. Большая часть желчных кислот конъюгирована с глицином или таурином. По химической природе желчные кислоты являются производными холановой кислоты
В желчи в основном содержится холевая, дезоксихолевая и хенодезоксихолевая кислоты

Считают, что только комбинация соль желчной кислоты + ненасыщенная жирная кислота + моноглицерид придает необходимую степень эмульгирования жира. Соли желчных кислот резко уменьшают поверхностное натяжение на поверхности раздела жир/вода, благодаря чему они не только облегчают эмульгирование, но и стабилизируют уже образовавшуюся эмульсию.
Основное расщепление липидов происходит в кишечнике, в первую очередь в двенадцатиперстной кишке. В этот отдел кишечника поступает сок поджелудочной железы, содержащий очень активную липазу. Сюда же поступает из желчного пузыря желчь, составные компоненты которой (желчные кислоты) необходимы для переваривания липидов. Это связано с тем, что желчные кислоты—холевая (преобладает в желчи человека), дезоксихолевая, литохолевая, хенодезоксихолевая, таурохолевая и гликохолевая —представляют собой поверхностно-активные вещества, способствующие эмульгированию жиров, что является важнейшим условием их последующего ферментативного расщепления.
Пройдя через барьер слизистой оболочки кишечника, желчные кислоты в связанном состоянии с липидами отделяются от последних и по венам кишечника через портальный кровоток возвращаются в печень, а затем с желчью в двенадцатиперстную кишку.
Образование эмульсии жиров в кишечнике может происходить и под влиянием мелких пузырьков СО2, выделяющегося при нейтрализации соляной кислоты пищевой кашицы бикарбонатами поджелудочного и кишечного сока. Способствуют эмульгированию и соли жирных кислот (мыла), возникающие при гидролизе липидов. Но основная роль в эмульгировании жиров принадлежит желчным кислотам.
В результате описанных процессов образуется очень тонкая жировая эмульсия, диаметр частиц которой не превышает 0,5 мкм. Такие эмульгированные жиры способны самостоятельно проходить через стенку кишечника и попадать в лимфатическую систему. Однако большая часть эмульгированного жира всасывается после гидролитического расщепления его панкреатическими липазами. Последние образуются в поджелудочной железе в виде неактивных проферментов, которые переходят в активную форму при участии мыльных кислот.
Основная масса липидов пищи представлена триацилглицеринами, меньше фосфолипидами и стероидами. Гидролиз триацилглицеринов идет постепенно. Сначала расщепляются эфирные связи в I м и 3-м положениях, т.е. внешние сложноэфирные связи:
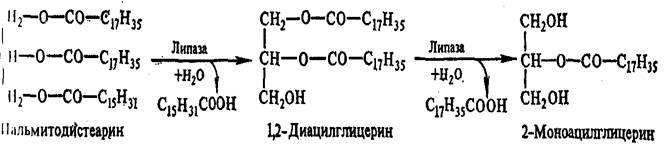
Эти реакции осуществляют липазы, специфичные в отношении 1,3-эфирных связей триацилглицерина. Связи во 2-м положении гидролизуют другие липазы:
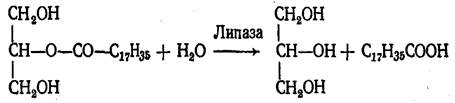
Связи 1 и 3 гидролизуются быстро, а потом идет медленный гидролиз 2-моноглицерида. 2-Моноглицерид может всасываться стенкой кишечника и использоваться на ресинтез триацилглицеринов, специфичных для данного вида организмов, уже в самой слизистой тонкого кишечника.
Кроме липаз в соке поджелудочной железы присутствуют эстеразы, гидролизующие преимущественно эфиры жирных кислот с короткой цепью и эфиры холестерина. Эти эстеразы тоже активны только в присутствии желчных кислот.
Пищеварительные липазы кроме человека и млекопитающих животных обнаружены и исследованы у рыб, некоторых беспозвоночных. Однако, как правило, у большинства видов беспозвоночных и костистых рыб липолитическая активность в пищеварительных соках примерно в 1000 раз ниже, чем в панкреатическом соке млекопитающих. Не следует забывать, что жиры могут усваиваться также путем фагоцитоза и сохраняться без предварительного гидролиза до тех пор, пока не прогидролизуются внутриклеточными липазами и, таким образом, примут участие в синтезе липидов в процессах образования энергии.
Расщепление фосфолипидов происходит при участии ряда ферментов: фосфолипаз А1, А2, С, D и лизофосфолипазы.
Фосфолипаза А1 гидролизует связь в 1-м положении. Фосфолипаза А2, образующаяся в поджелудочной железе, поступает в полость тонкого кишечника в неактивной форме и только под действием трипсина активируется. Под действием фосфорилапазы А2 отщепляется жирная кислота во 2-м положении. В результате ее действия образуются лизофосфолипиды, которые вызывают разрушение триглицеридов крови. Кроме панкреатического сока фосфолипаза А2 содержится в яде рептилий, беспозвоночных (особенно членистоногих — пчел, скорпионов, муравьев), а также у кишечнополостных. Известны так|же внутриклеточные фосфолипазы А2 (в лизосомах, микросомах, митохондриях).
В организме ее действие компенсируется фосфорилазой А1, которая отщепляет второй кислотный остаток. Затем отщепляется азотистое основание под действием фосфорилазы D и фосфорная кислота – фосфорилазой С.
Конечными продуктами распада фосфолипидов являются жирные кислоты, глицерин, азотистое основание и фосфорная кислота.
Стериды, подвергаясь действию гидролитических ферментов типа холестераз, расщепляются в кишечнике с образованием спирта холестерола или эргостерола и соответствующей жирной кислоты. Холестеразы продуцируются поджелудочной железой и активны только в присутствии солей желчных кислот.
Таким образом, образующаяся в результате гидролиза липидов смесь содержит анионы жирных кислот, моно-, ди- и триацилглицерины, хорошо эмульгированные солями жирных кислот и мылами, глицерин, холин, этаноламин и другие полярные компоненты липидов. Исследования с мечеными триацилглицеринами показали, что около 40% жиров пищи гидролизуется полностью до глицерина и жирных кислот, 3—10% всасываются без гидролиза в форме триацилглицеринов, а остальные гидролизуются частично, главным образом до 2-моноацилглицеринов. Глицерин водорастворим и вместе с жирными кислотами, имеющими короткие углеродные цепи (С<10), всасывается свободно через стенку кишечника и через портальную систему кровообращения поступает в печень.
Для всасывания жирных кислот с длинной цепью (С >10), моноглицеридов и холестерина необходимы желчные кислоты. Соединяясь с вышеперечисленными соединениями, желчные кислоты образуют растворимые комплексы или мицеллы— холеиновые комплексы, которые легко всасываются в эпителий кишечника. Так как рН в тонком кишечнике слабощелочная, желчные кислоты функционируют здесь в форме своих солей. Особую роль при этом играют такие желчные кислоты, как таурохолевая и гликохолевая. Лучше перевариваются и всасываются липиды, находящиеся в жидком состоянии, при температуре тела. Липиды, у которых точка плавления существенно выше температуры тела, плохо перевариваются и всасываются.
Фосфорная кислота, образующаяся при гидролизе фосфолипидов, всасывается в виде натриевых и калиевых солей, а азотистые основания — холин, этаноламин и серин — всасываются при участии нуклеотидов (ЦДФ-производных). Некоторая избирательность проявляется слизистой оболочкой кишечника в отношении стероидов, особенно растительного происхождения. Среди основных стероидов пищи только холестерин легко проникает через стенки кишечника. С такой же легкостью всасываются витамин D и некоторые стероидные гормоны, введенные перорально.
Преобладающими липидами лимфы являются триацилглицериды, даже тогда, когда жирные кислоты находятся в составе сложных эфиров других спиртов.
Желчные кислоты выполняют в организме 3 основные функции:эмульгируют жиры;активируют липазу;обеспечивают всасывание высших жирных кислот, моноглицеридов и холестерина.
.
Date: 2015-05-22; view: 662; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |