Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Генетическая структура анализируемых групп КРС по гену бета-лактоглобулина определенная по методикам Гладырь и др., 2001 и Medrano, et al., 1900
| порода | n | Частота аллелей и генотипов, % по методике Гладырь и др.,2001 | Частота аллелей и генотипов, % по методике Medrano, et al., 1900 | Частота несовпадений генотипов, % | ||||||||||||
| АA | AВ | ВB | А | В | АA | AВ | ВВ | А | В | |||||||
| Черно-пестрая | 19,4 | 30,5 | 0,41 | 0,58 | 19,4 | 47,2 | 33,3 | 0,43 | 0,57 | 2,7 | ||||||
| Hex | 0,493 | 0,49 | ||||||||||||||
| Hobs | 0,5 | 0,472 | ||||||||||||||
| χ2 | 0,0043 | |||||||||||||||
| Айрширская | 6,6 | 31,3 | 62,2 | 0,22 | 0,77 | 2,2 | 26,6 | 71,1 | 0,15 | 0,84 | 13,3 | |||||
| Hex | 0,345 | 0,262 | ||||||||||||||
| Hobs | 0,311 | 0,266 | ||||||||||||||
| χ2 | 0,8445 | - | ||||||||||||||
| Частный скот | 21,3 | 52,4 | 26,2 | 0,47 | 0,52 | 18,0 | 42,6 | 39,3 | 0,39 | 0,6 | 9,7 | |||||
| Hex | 0,498 | 0,477 | ||||||||||||||
| Hobs | 0,524 | 0,426 | ||||||||||||||
| χ2 | 0,3963 | - | ||||||||||||||

Рис.3.6.3- Соотношение частот генотипов β-лактоглобулина в анализируемых группах КРС. 1 – группа коров черно-пестрой породы, ОАО «Снежка-Госома; 2 – частный скот (Жирятинский р-н, Брянской области); 3 – группа коров айрширской породы, СПХ Сельцо.
В таблице 3.6.2 приведены частоты А-аллеля β-лактоглобулина в группах крупного рогатого скота разных пород. Из таблицы видно, что преобладание В-аллеля наблюдается в разных регионах мира. Исключением являются некоторые группы коров голштинской и чешской красной. В этих группах частота А-аллеля превышает частоту В-аллеля. Кроме того, для разных групп крупного рогатого скота голштинской породы из разных регионов мира в литературе приводятся близкие значения частот аллелей. Так, например, в группе голштинов из США, Ирана, Мексики, Украины, Аргентины частоты А-аллеля составляют - 0,530, 0529, 0,470, 0460, 0,43 соответственно. Вероятно, что для голштинской породы частота аллелей гена бета-лактоглобулина может является породоспецифичной характеристикой. Ранее, Глазко ранее отмечал у голштинов по локусу каппа-казеина незначительные межгрупповые различия, не зависящие от региона разведения [Глазко, 2006].
Частота А-аллеля в исследуемых нами группах частного скота и коров черно-пестрой породы приближенно соответствует частотам этого аллеля в разных группах КРС голштинской породы, что косвенно может быть связано с голштинизацией.
Таблица 3.6.2
Частота А-аллеля бета-лактоглобулина в группах КРС Брянской области и в других регионах
| порода | Частота А-аллеля | Автор | Регион |
| Чешская красная | 0,571 | M Zatoń-Dobrowolskaet al., 2006 | Польша |
| Caracu | 0,57 | Kemenes et al.1999 | Бразилия |
| Голштинская | 0,530 | A. Barreras et al., 2001 | США |
| Голштинская | 0,529 | Heidari et al. | Иран |
| Якутская | 0,516 | Гладырь и др., 2000 | Россия |
| Черно-пестрая | 0,48 | Гладырь и др., 2000 | Россия |
| Частный скот | 0,47 | Собственные данные | Россия, Брянск |
| Голштинская | 0,470 | А. Barreras et al., 2002 | Мексика |
| Голштинская | 0,460 | Копылова К.В., и др., 2009 | Украина |
| Romanian Spotted | 0,450 | Ilie D et al., 2008 | Румыния |
| Голштинская | 0,43 | A. G. Bonvillani et al., 1998 | Аргентина |
| Черно-пестрая | 0,41 | Собственные данные | Россия, Брянск |
| Швицкая | 0,406 | Гладырь и др., 2000 | Россия |
| Nelore (Bos indicus) | 0,40 | Kemenes et al.1999 | Бразилия |
| Польская красная | 0,397 | M Zatoń-Dobrowolskaet al., 2006 | Польша |
| Tharparkar(Bos indicus) | 0,39 | S. Rachagani et al., 2006 | Индия |
| Gyr | 0,37 | Kemenes et al.1999 | Бразилия |
| Brown of Maramures | 0,333 | Ilie D et al., 2008 | Румыния |
| Эстонская голштинская | 0,319 | Sirje Varv, et al., 2009 | Эстония |
| Bubalus bubalis | 0,301 | Rajesh K et al., 2007 | Индия |
| Jersey × Zebu, | 0,30 | Rajesh K. PATEL et al., 2007 | Индия |
| Украинская черно-пестрая | 0,278 | Малиенко В.А., и др., 2008 | Украина |
| Голштино-фризская × Zebu | 0,26 | Rajesh K. PATEL et al., 2007 | Индия |
| Красная горбатовская | 0,246 | Гладырь и др., 2000 | Россия |
| Айрширская | 0,22 | Собственные данные | Россия, Брянск |
| Hanwoo | 0,208 | Kim, J et al., 1998 | Корея |
| Германская красная | 0,191 | M Zatoń-Dobrowolskaet al., 2006 | Польша |
| Эстонская | 0,188 | Sirje Varv, et al., 2009 | Эстония |
| Sahiwal (Bos indicus) | 0,17 | S. Rachagani et al., 2006 | Индия |
| Украинская черно-пестрая | 0,165 | Копылова К.В., и др., 2009 | Украина |
| Корейская | 0,153 | Chung, E. R et al., 1998 | Корея |
| Эстонская красная | 0,138 | Sirje Varv, et al., 2009 | Эстония |
| Датская красная | 0,110 | M Zatoń-Dobrowolskaet al., 2006 | Польша |
| Абердин-ангусская | 0,2 | Копылова К.В., и др., 2009 | Украина |
3.7. Анализ сочетания мутаций при определении А и В аллелей гена β-лактоглобулина.
В связи с тем, что в настоящее время разработаны две методики, позволяющие согласованно выявлять А и В варианты гена бета-лактоглобулина, стало возможным предположить наличие рекомбинации между мутациями расположенных в разных частях гена типирующих В-аллель βLG. в основе выбранных нами методик лежит анализ точечных нуклеотидных замен в экзонах 2 и 4. Каждая из этих методик выявляет одну из двух мутаций характерных для В-аллеля. Проводя одновременно анализ по этим методикам можно учитывать эти две замены.
При анализе полиморфизма гена бета-лактоглобулина мы обнаружили несоответствия в определении аллелей этого гена по описанным методикам. На рисунке 3.7.1. представлена электрофореграмма, которая иллюстрирует несовпадения при определении аллельных вариантов по представленным методикам. Особенно интересен результат рестрикции фрагментов экзонов 2 и 4 полученных на ДНК образца 109. При рестрикции фрагмента второго экзона 1247 п.о. эндонуклеазой PvuII выявляется типичная картина характерная для АА – варианта гена бета-лактоглобулина, т.е. отсутствие нуклеотидной замены, обеспечивающей появление дополнительного сайта для PvuII, описанного для ВВ-варианта. Напротив, при рестрикции фрагмента экзона 4 этого же образца эндонуклеазой HaeIII наблюдается появление специфичных для В-аллеля фрагментов.
Частоты несовпадении при определении генотипов по двум вышеописанным методикам в группах коров черно-пестрой, айрширской пород и частного скота составили – 2,7%, 13,3%, 9,7% соответственно.
В таблице 3.7.1. отображены частоты встреченных сочетаний аллельных вариантов при определении полиморфизма гена βLG по методикам Гладырь и Medrano [Гладырь и др., 2001 и Medrano, et al., 1990]. С наивысшей частотой встречались сочетания АВ/ВВ (0,61) и АА/ВВ (0,22). Сочетание ВВ/АВ было самым редким, а сочетания АВ/АА и ВВ/АА не обнаружены. Генотипы АВ/АВ неотличимы от обычных гетерозигот АВ.
Таблица 3.7.1
Частоты сочетаний мутаций в генотипах КРС при определении полиморфизма гена бета-лактоглобулина по описанным методикам.
| Варианты гена βLG по методике Гладырь и др., 2001 г. | Варианты гена βLG по методике Medrano, et al.,1990 | ||
| ВВ | АВ | АА | |
| АВ | 0.61 | - | |
| АА | 0.22 | 0.11 | |
| ВВ | 0.055 | - |
На рисунке 3.7.2 представлена схема, раскрывающая филогенетические связи между разными вариантами гена бета-лактоглобулина [Formaggioni P., 1999]. В основе филогенетического дерева авторы расположили В-аллель βLG. Добавляя к этому варианту определенные мутации можно прийти ко всем остальным аллелям.
Проведенные нами исследования по выявлению двух мутаций определяющих В-вариант в целом соответствуют приведенной филогенетической связи А- и В-аллелей, но выявленные нами сочетания мутаций не вкладываются в эту схему. Руководствуясь результатами наших исследований, можно предположить, что существуют промежуточные между А- и В-аллелями варианты или, что произошла рекомбинация между мутациями во втором и четвертом экзонах гена бета-лактоглобулина с образованием новых аллелей (Рис. 3.7.3.).
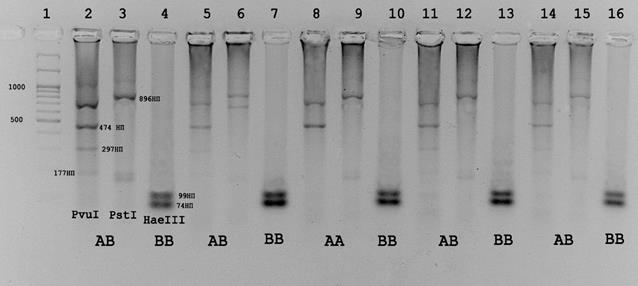
Рис.3.7.1 - Электрофореграмма продуктов рестрикции при определении вариантов гена бета-лактоглобулина по методикам Гладырь и др., 2001 и Medrano, et al., 1990. 1 – маркер молекулярных масс М27(СибЭнзим). 2,3,4-анализ образца 23; 5, 6, 7 – образец 99; 8, 9, 10 – образец 109; 11, 12, 13 - образец 96; 14, 15, 16 – образец 181. Каждый первый из тройки трек – рестрикция эндонуклеазой PvuII, каждый второй - продукты рестрикции PstI, каждый третий - продукты рестрикции HaeIII.

Рис. 3.7.2 - Схема происхождениягенетических вариантов гена бета-лактоглобулина гена бета-лактоглобулина (по P. Formaggioni et all.,1999)
 |

Рис. 3.7.3 - Вероятное положение промежуточных вариантов между А- и В-аллелями в схеме филогении полиморфизма гена бета-лактоглобулина.
Date: 2015-11-14; view: 478; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |