Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Эволюция уха позвоночных
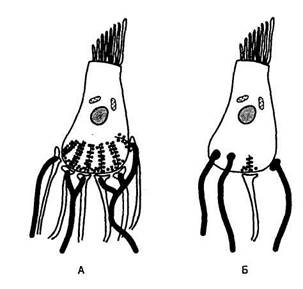
Несомненно, система боковой линии и ухо тесно связаны. У миног впячива-ние на голове быстро изолируется от внешней среды и развивается в две заполненных жидкостью трубки под прямым углом друг к другу и две камеры, также заполненных жидкостью. Трубки — это полукружные каналы, а камеры — утрикулюс (маточка) и саккулюс (мешочек). Вместе они образуют мембранное преддверие. Многие рассматривают его как специализированный отдел системы боковой линии в голове, а его организация лежит в основе, как показывает рис. 8.8, структуры уха всех позвоночных. Только у крокодилов, а также птиц и млекопитающих, неприметный у рыб вырост саккулюса — лагена (lagaena), претерпевает поразительное развитие и образует собственно улитку (cochlea). Вместе полукружные каналы, утрикулюс, саккулюс и лагена (или улитка птиц и млекопитающих) образуют мембранный лабиринт.
8.3.1. Равновесие
Ухо первоначально было органом баланса и равновесия. Детектирование звука, которое принимает самодовлеющие формы в мире млекопитающих, и людей в частности — это позднейшее добавление. Мембранное преддверие осталось поразительно неизменным за те полмиллиарда лет, что прошли с возникновения позвоночных (рис. 8.8). У всех челюстноротых позвоночных добавился еще один полукружный канал, ортогональный тем, которые присутствуют у миноги. Вся система заполнена водным раствором — эндолимфой — и подвешена в полости уха, где она плавает в другом водном растворе — перилимфе. Эти две жидкости радикально различаются по ионному составу. Тогда как пе-рилимфа напоминает другие внеклеточные жидкости и имеет высокую концентрацию Na+ (150 мМ/л) и низкую — К+ (3-4 мМ/л), эндолимфа намного более схожа с внутриклеточной средой, будучи богатой К+ (150 мМ/л) и бед-

Рис. 8.8. Эволюция мембранного лабиринта. (А) минога Petromyzon; (Б) акула Scyllium; (В) макрель Scomber, (Г) лягушка Rana; (Д) ящерица Lacerta; (Е) голубь Columba; (Ж) морская свинка Cavia. Для рисунков А, Б, В, Г, Е, Ж: ап — ампула переднего канала, аг — ампула горизонтального канала, аз — ампула заднего канала, пвк — передний вертикальный канал, у — проток улитки, ее — cms commune, общая ножка, к — камеры, выстланные ресничным эпителием; эп — эвдолимфатический проток (ductus endolynphaticus), гк — горизонтальный канал, 1 — lagaena, мл — макула улитки lagaena, мн — macula neglecta (макула незаметная), мс — макула саккулюса, му — макула утрикулюса, ко — кортиев орган, ра — papilla amphibiorum, pb — papilla basilaris, звк — задний вентральный канал, с — саккулюс, у — утрикулюс. Для рисунка Д: ппк — передний полукружный канал, pb —papilla basilaris, ее — cms commune, пу — проток улитки, мл — макула улитки lagaena, лпк — латеральный полукружный канал, мн — macula neglecta, сп — слуховой проход, зпк — задний полукружный канал, мс — макула саккулюса, у — утрикулюс, му — макула утрикулюса, yen — утрикуло-саккулярный проток. Части рисунка А, Б, В, Г, Е и Ж — из Romer, 1970. Воспроизводится с разрешения W. В. Saunders Company; часть рисунка Д — из Baird, 1974. Воспроизводится с разрешения Springer-Verlag GmbH & Co. KG.
ной Na+ (1—2,5 мМ/л). Выше мы отмечали, обсуждая биофизику волосковых клеток, что ионные потоки в стереоцилии, главным образом, — это потоки К+. Ниже мы увидим, что стереоцилии простираются в эндолимфу, а необычный ионный состав эндолимфы играет ключевую роль в физиологии внутреннего уха. Наконец, отметим, что, хотя лабиринт свободно плавает в полости уха, он подвешен к стенкам полости коллагеновыми волокнами. В большинстве случаев тонкая трубка эндолимфатического протока {ductus endolymphaticus) простирается от саккулюса до полости черепа, где оканчивается эндолимфатиче-ским мешком.
В стенках у утрикулюса и саккулюса располагаются «пятна», содержащие сенсорные волосковые клетки — макулы саккулюса и утрикулюса. Макула утрикулюса лежит в дне компартмента, а макула саккулюса — обычно в вертикальной плоскости — в стенке камеры (рис. 8.9 А). Макулы иннервированы волокнами, принадлежащими вестибулярному нерву. У рыб есть небольшое удлиннение саккулюса — лагена, тоже содержащее сенсорную макулу. Волосковые клетки макул очень похожи на клетки органов боковой линии. Здесь, однако, могут быть выделены 2 типа клеток: амфороподобные клетки типа 1 и цилиндрические клетки типа 2 (рис. 8.9 Б). Как и в органах боковой линии, волосковые клетки погружены в желатинозную купулу. В утрикулюсе, саккулю-се и лагене желатинозная купула часто импрегнирована кристаллами СаСО3, образующими отолиты или «слуховые камешки». Отолит саккулюса обычно развивается в наибольшей степени и у многих костистых рыб почти заполняет собой камеру. Форма отолитов у разных видов рыб настолько специфична, что может быть использована для определения вида.
Функции утрикулюса и саккулюса достаточно просты — это восприятие линейного ускорения головы. Возможны два случая. В первом голова движется вперед-назад или вверх-вниз — при этом инерция отолитов обеспечивает изгибание стереоцилии волосковых клеток в ту или иную сторону. Во втором случае, когда голова неподвижна, линейное ускорение отолитов, вызванное силой тяжести, заставляет их смещаться вниз относительно волосковых клеток. Сила тяжести действует на волосковые клетки в разных направлениях в зависимости от угла наклона головы, и эти различающиеся изгибы детектируются стереоцилиями описанным выше образом. Информация передается в мозг по вестибулярному нерву.
Три полукружных канала расположены в трех плоскостях, все они исходят из утрикулюса (рис. 8.8), и в каждом имеется сферическое расширение — ампула, через которую они сообщаются с этой камерой. Каналы, как и прочие части мембранного преддверия, заполнены эндолимфой. Внутри ампул расположены чувствительные участки — гребешки — в которых также присутствуют волосковые клетки. Кончики ресничек последних погружены в большую желатинозную купулу, которая почти полностью закрывает просвет ампулы, смещаясь подобно двери в метро — в ту и другую сторону.
Функция полукружных каналов — это детектирование угловых ускорений головы. Когда голова вращается в любой из трех плоскостей, инерция эндолимфы создает некоторую задержку сдвига. Затем взаимодействие ее вязких свойств со стенкой полукружного канала приводит ее в движение, и далее она движется в унисон в содержащей ее структурой. Однако, первичная задержка

|
Рис. 8.9. (А) Расположение волосковых клеток в (i) ут-рикулюсе и (ii) саккулюсе и лагене. (Б) Два типа волосковых клеток в макулах и гребнях мембранного лабиринта: амфороподобные клетки типа 1 и цилиндрические клетки типа 2. Клетки типа 1 окружены чашевидными нервными окончаниями. В правом верхнем углу рисунка — срез через реснички клетки типа 2 по линии А—А. Киноцилия обозначена черным кружком, эо — эфферентное нервное окончание, ос — область синапса, нно — не-миелинизированное нервное окончание. Воспроизводится с разрешения A. J. Benson.
движения эндолимфы по отношению к содержащему ее протоку означает, что «дверь» гребешка ампулы открывается. Другими словами, желатинозная купу-ла и погруженные в нее стереоцилии изгибаются, что и запускает импульс в волокнах вестибулярного нерва. Отметьте, что детектируемые угловые ускорения — это не постоянные скорости, которые могут быть вызваны длительным вращением (порядка 20 с).
Сенсорная информация от утрикулюса, саккулюса и полукружных каналов интегрируется с информацией, получаемой глазами, от рецепторов мышц, связок и кожи и инициирует рефлексы, поддерживающие нормальную ориентацию животного по отношению к вектору силы тяжести и противодействующие приложенным внешним ускорениям во всех плоскостях. Большинство из этих рефлексов опосредованы спинным мозгом и стволом головного мозга, кора в них задействована очень в небольшой степени.
ухо у птиц состоит из трех отделов: наружного слухового прохода, ведущего к барабанной перепонке, среднего уха, пересеченного слуховым стержнем и тремя неправильной формы косточками, и внутреннего уха, содержащего мембранный лабиринт.
Большинство птиц, естественно, чрезвычайно чувствительны к звуку, который играет огромную роль в их социальном поведении. Песни птиц видо-специфичны, причем ясно, что птенцы выучивают ее локальный тип. Выяснилось, что некоторые из птиц, обитающих в пещерах, используют звуки и для эхолокации {Steatornis — венесуэльский гуахаро, Collacalia — азиатский стриж), однако нет никаких данных, которые бы свидетельствовали о том, что у птиц развита чувствительность к ультразвуковым колебаниям, которая характерна для летучих мышей. Наконец, чувствительность к направлению звука достигает очень высокого уровня у ночных хищников, таких как совы (см. раздел 9.3).
i/Млекопитающие
У млекопитающих слуховая часть уха — улитка — достигает эволюционной вершины. Самый слабый звук, который способен уловить человек, соответствует около 0,0001 дин/см2 или прибл. 10~16 Вт/см2. Это соответствует изменению давления на барабанной перепонке в 20 мкПа. Диапазон частот, которые способен ощущать человек, простирается от 20 Гц до 20 кГц, хотя с возрастом верхняя граница чувствительности снижается. Тренированное ухо может различать частоты 1000 и 1002 Гц. Такая поразительная способность — следствие высокого развития улитки. Как показывает рис. 8.8 Ж, у млекопитающих улитка уже не прямая или слабо изогнутая, как у птиц, а свернута в спираль, похожую по форме на раковину моллюска. Срез ее показан на рис. 8.13 А. Из рисунка видно, что три канала — вестибулярная, средняя и барабанная лестницы, с которыми мы впервые встречались у пресмыкающихся, присутствуют и здесь, а рис. 8.13 Б показывает, что базилярный сосочек развивается здесь в сложный кортиев орган.
Прежде чем мы сможем обсудить эту сложную тонкую структуру, необходимо сделать краткий обзор уха млекопитающих в целом. Рис. 8.14 А показывает, что и наружное и среднее ухо претерпели значительное развитие. У многих видов ушная раковина (не показана) велика и достаточно подвижна. Это играет значительную роль в определении направления на источник звука. У людей мышцы, двигающие ушную раковину у других животных, сохраняются в основном в рудиментарной форме. Наружный слуховой проход сравнительно длинный и, для предупреждения нежелательных посетителей, покрыт волосами и железами, секретирующими ушную серу. Барабанная перепонка представляет собой границу между наружным и средним ухом. В среднем ухе поддерживается атмосферное давление за счет прохода через евстахиеву трубу, открывающуюся в глотку. Это важно, поскольку барабанная перепонка тонка, и резкое различие давления по обе ее стороны может привести к ее разрыву. Вибрация барабанной перепонки, вызываемая звуком, передается через камеру среднего уха слуховыми косточками: молоточком (malleus), наковальней (incus) и стременем (stapes). Эти, ставшие притчей во языцех, самые маленькие

Рис. 8.13. (А) Канал улитки изогнут в форме раковины моллюска. Срез на рисунке проходит вертикально через улитку. (Б) Средняя лестница и кортиев орган — более детально. Из Freeman and Bracegirdle, 1976. Воспроизводится с разрешения Butterworth Heinemann.
кости в организме человека прикреплены к стенке камеры среднего уха связками и мышцами. Мышцы, т. tensor tympani, идущая к молоточку, и т. stapedius, идущая к стремечку, сокращаются одновременно в ответ на звуки высокой интенсивности и таким образом защищают тонкие структуры внутреннего уха от повреждающего эффекта избыточной вибрации. Наконец, основание стремечка, овальная пластинка (basis stapedius) сочленяется с мембраной, закрывающей овальное окно (fenestra ovalis) круговой связкой. При развитии отосклероза эта часть стремени срастается с костным окружением овального окна, что ведет к глухоте.
Теперь вернемся к физиологии улитки. Для упрощения рис. 8.14 Б показывает улитку, развернутой из спирали в прямую. Мы уже видели, что, когда вол-

Рис. 8.14. (А) Анатомия уха млекопитающего. (Б) Принципиальная схема улитки, развернутой в прямую. Детальные объяснения в тексте. Из Smith, 1970, с разрешения.
на атмосферного давления, которую мы воспринимаем как звук, достигает барабанной перепонкой, вибрация передается в овальное окно. Таким образом, мембрана, покрывающая окно, передает изменения давления перилимфе внутреннего уха. Теперь рассмотрим рис. 8.13 Б и 8.14 Б. Поскольку мембрана, покрывающая овальное окно смещается внутрь, давление в вестибулярной лестнице возрастает. Рейснерова мембрана сдвигается вниз, что увеличивает давление в средней лестнице до тех пор, пока оно не компенсируется смещением базилярной мембраны. Возникающее вследствие этого избыточное давление в барабанной лестнице компенсируется изгибанием наружу мембраны, покрывающей круглое окно (fenestra rotunda). Все эти движения, конечно, происходят практически мгновенно.
Как видно из рис. 8.13 Б, в рассматриваемой системе присутствует еще одна — текториальная мембрана, опирающаяся на два ряда волосковых клеток. Кончики ресничек погружены в нее, точно также, как в описанном выше случае — в желатинозную купулу. В отличие от рейснеровой и основной, тектори-

Рис. 8.15. Сканирующая электронная микрофотография волосковых клеток кортиева органа. Текториальная мембрана удалена, а микроскоп направлен на основную мембрану. (А) Три ряда наружных волосковых клеток (отметьте U-образное расположение) и один ряд внутренних волосковых клеток. (Б) стереоцилии внутренних волосковых клеток образуют почти прямую линию. (В) Внутренние волосковые клетки при большем увеличением (7500х). (Г) Стереоцилии наружных волосковых клеток меньше и выстроены в форме буквы U. (Д) Стереоцилии наружных волосковых клеток при большем увеличении (12 500х). (А), (Б) и (Г) из Pickles, 1988; воспроизводится с разрешения Academic Press Ltd. (В) и (Д) — Pickles, личное сообщение, воспроизводится с разрешения.
альная мембрана сравнительно свободно плавает в эндолимфе и следовательно изменения гидростатического давления влияют на нее в равной степени со всех сторон. Поэтому изменения давления, особенно быстрые изменения, не могут заставить эту мембрану двигаться вверх или вниз, синхронно с другими двумя.
Теперь рассмотрим волосковые клетки. Рис. 8.13 Б показывает, что здесь присутствует один ряд внутренних волосковых клеток, отделенных небольшим пространством от трех или четырех рядов наружных волосковых клеток. У этих клеток развивается от 50 до 100 стереоцилий, однако, в отличие от уже рассмотренных волосковых клеток, здесь нет киноцилий. На том месте, где можно было бы ожидать присутствия киноцилий, развивается крупная цент-риоль. Вид клетки снаружи показывает, что стереоцилий принимают форму буквы «W» или «U», нижние концы которых направлены к центриоли (рис. 8.15). Кончики стереоцилий погружены в текториальную мембрану, а основания волосковых клеток присоединены в базилярной мембране, где они образуют синаптические контакты с дендритическими окончаниями кохлеар-ного нерва.
Интересны эти синапсы волосковых клеток. Рис. 8.16 показывает множество синаптических везикул (вероятно, содержащих глютамат), концентрирующихся вокруг электронно-плотного материала, который, как и синаптические ленты в фоторецепторах позвоночных, служит для организации везикул перед их выбросом. Синапсы содержат различные механизмы в пресинаптиче-ской зоне, обеспечивающие быстрое и эффективное выделение медиатора. Здесь концентрируются Са2+-каналы такого типа, который быстро реагирует на малые изменения потенциала. Это обстоятельство обеспечивает и почти мгновенное прекращение выброса медиатора. Существуют еще два механизма быстрого прекращения выброса медиатора: цитоплазма волосковых клеток — очень жесткий кальциевый буфер, к тому же быстрая реполяризация мембраны обеспечивается за счет концентрации Са2+-зависимых К+-каналов. Все эти специализированные механизмы помогают предупредить потерю быстрого ответа на механическую деформацию пучка ресничек в медленном синаптиче-ском механизме в основании волосковых клеток.
Между внутренними и наружными волосковыми клетками есть существенное различие. Тогда как внутренние волосковые клетки образуют контакты с прибл. десятком афферентных волокон кохлеарного нерва, наружные много менее иннервированы. Около 90% афферентных волокон кохлеарного нерва идет от синаптических окончаний на ряде внутренних волосковых клеток. Рис. 8.16 показывает, что основания волосковых клеток содержат множество синаптических пузырьков (везикул), а также «электронно-плотное тело» (синаптические ленты) и другие дополнительные структуры. Присутствуют также эфферентные волокна кохлеарного нерва. Рис. 8.16 также показывает, что эти волокна образуют синаптические контакты с дендритными окончаниями афферентных волокон. Из этого можно предположить, что здесь над чувствительностью системы существует контроль обратных связей. Наружные волосковые клетки, напротив, бедно иннервированные афферентными волокнами, получают мощную эфферентную иннервацию. Окончания эфферентных волокон — крупные, заполненные медиаторными везикулами. В информацион-
ном дополнении 8.1 мы увидим, что наружные волосковые клетки обладают сократительной способностью. Вероятно, такая эфферентная иннервация также позволяет мозгу контролировать чувствительность улитки путем изменения расстояния между текториальной и базилярной мембранами (мы вернемся к этому вопросу, когда будем рассматривать физиологию волокон улитки в гл. 9).
Как и можно было ожидать от такого тонкого и точного механизма, многое в нем может ломаться и приводить к глухоте. С огромным ростом в последние годы знаний о человеческом геноме открыто более сотни «генов глухоты». Они расположены во всех 22 аутосомах, а также в Х-хромосоме. Многие из них связаны с другими (не слуховыми) аномалиями, однако, по крайней мере дюжина стоят особняком и не являются синдромными, хотя какие-то другие признаки и могут быть в конце концов связаны с ними. Конечно, далеко не вся сотня генов ответственна за формирование уха и волосковых клеток. Многие из них в большей степени влияют на развитие центральных отделов слуховой системы, однако те, которые отвечают именно за волосковые клетки, позволяют начать формирование генетического понимания этих жизненно важных структур. Это например ген, кодирующий калиевый канал в наружных волосковых клетках; гены коннексинов, значительная роль которых в калиевом го-меостазе также показана, и открытый сравнительно недавно ген OTOF в хромосоме 2, который кодирует белок отоферлин, экпрессирующийся во внутренних волосковых клетках. Существуют доказательства того, что отоферлин вовлечен в выстраивание везикул вдоль синаптических лент, характерных, как мы видели выше, для синапсов волосковых клеток.
|
Рис. 8.16. Иннервация внутренних и наружных волосковых клеток кортиева органа. Схематический рисунок показывает афферентные (белые) и эфферентные (черные) волокна. (А) внутренняя волосковая клетка. Эфферентные волокна образуют синаптический контакт с дендритными окончаниями афферентных волокон. (Б) Наружная волосковая клетка. Эфферентные волокна образуют синапсы непосредственно на во-лосковой клетке, которая имеет лишь небольшое число синапсов (показан только один) с сенсорными (афферентными) волокнами.
Теперь обратим внимание на основную тему данного предмета. Мы видели, что базилярная мембрана колеблется в ответ на поступающий в ухо звук, тогда как текториальная мембрана остается сравнительно неподвижной. Сте-реоцилии волосковых клеток подвергаются механической деформации, причем их реснички погружены в богатую К+ эндолимфу. Возникающая деполяризация может быть обнаружена с помощью микроэлектродных отведений. Они точно воспроизводят частоту поступающего звука. Это т. н. микрофонные потенциалы. Микрофонные деполяризации (рецепторные потенциалы) ведут к выделению медиаторных веществ на дендритные окончания афферентных волокон кохлеарного нерва.
Таким образом, мы видим, что в самой основе потрясающе сложного внутреннего уха млекопитающих лежат волосковые клетки; конечно, модифицированные, но в целом те же самые, что мы впервые встретили в каналах органа боковой линии наших водных предшественников. Мы еще увидим, что примерно то же самое можно сказать и о других органах чувств. Молекулярные механизмы, развившиеся в эволюционной истории очень рано, сохраняются, но со временем оказываются встроенными в невероятно сложные и хитроумные органы.
Одним из эволюционных императивов, который двигал развитием улитки млекопитающих, была необходимость отличать разные частоты звука. Мы видели, что эта способность в небольшой степени присутствует у рыб, земноводных и пресмыкающихся; у птиц и млекопитающих она претерпевает огромное развитие. Выше мы упоминали, что частотный диапазон человеческого уха лежит между 20 Гц и 20 кГц (с некоторым снижением верхнего предела с возрастом). Мы также отмечали, что в пределах диапазона слышимости человек и другие млекопитающие обладают чрезвычайно высокой способностью к различению частот. А потому следующий вопрос — как она достигается?

|
Рис. 8.17. Схематическое изображение базилярной мембраны. Мембрана распрямлена. У человека они имеет около 33 мм в длину, а ширина меняется от прибл. 100 мкм у круглого окна до 500 мкм у геликотремы. Дальнейшие объяснения — в тексте.
Может показаться, что у этой проблемы — простое решение. Почему бы кохлеарному нерву не быть фазово-синхронным с приходящей звуковой волной давления? Иными словами, почему бы о тональной частоте 20 Гц не сигнализировать нервными импульсами частотой 20 Гц, а о тональной частоте 15 или 20 кГц — импульсами частотой 15 и 20 кГц, соответственно? В таком простом решении есть две очевидные трудности. Во-первых, как мы отмечали в гл. 2, частота импульсов в сенсорных нервах обычно сигнализирует об интенсивности стимула. Нервная система могла бы, конечно, обойти это затруднение, однако, вторая трудность более непреодолима. Биофизика нервных волокон такова, что за каждым импульсом следует рефрактерный период длительностью около 2 мс. Из этого вытекает (как мы видели в гл. 2), что одиночное волокно не способно проводить более 500 импульсов в секунду. Т. о., для частот выше 500 Гц нужны какие-то иные средства частотной дискриминации.
Здесь могут работать два основных механизма. Во-первых, есть данные (см. гл. 9), что кохлеарные волокна могут быть фазо-синхронными звуковым частотам выше 500 Гц, но не реагируя на каждый частотный импульс. То есть предполагается, что в нижней части частотного спектра (ниже 5 кГц) группа волокон кохлеарного нерва объединяется для достижения частоты импульсов, совпадающей с тональной частотой, в каком-то слуховом центре мозга. По понятным причинам, такая идея именуется теорией залпа. Второй, значительно более важный механизм основывается на наблюдении, что ширина базилярной мембраны увеличивается от круглого окна к геликотреме (или в случае птиц —к макуле улитки). Ширина базилярной мембраны человека, например, увеличивается с 100 до 500 мкм на расстоянии в 33 мм (рис. 8.17). Герман фон Гельмгольц еще в XIX веке предположил, что базилярную мембрану можно уподобить ряду настроенных камертонов (резонаторов). Тоны высокой частоты вызывают максимальные возмущения в области круглого окна, а низкой — у геликотремы. Точные исследования фон Бекеши (von Bekesy) и других в основном подтвердили гипотезу Гельмгольца. Обнаружено, что волны сложной формы двигаются вдоль всей базилярной мембраны, но место, где они достигают максимальной амплитуды, как и предположил Гельмгольц, связано с их частотой. Догадка Гельмгольца по очевидным причинам известна как теория
места частотной дискриминации. Чтобы различить частоты, мозгу достаточно лишь «посмотреть», из какого места базилярной мембраны происходят волокна, в которых активность максимальна.
В последнее время теория места Гельмгольца была расширена и подкреплена обнаружением того факта, что сами по себе волосковые клетки настроены на определенную частоту звука. Такая настройка — результат молекулярной анатомии и цитологии клетки. Показано, что волосковые клетки и их стереоцилии различаются в размерах и гибкости в разных частях базилярной мембраны. Вблизи от круглого окна стереоцилии малы и сравнительно жестки, тогда как в области геликотремы больше в размерах и гибки. Это вместе со специфической популяцией ионных каналов определяет электрические характеристики данной клетки (см. Информацию 8.1). Каждая волосковая клетка настроена на максимальную реакцию в ответ на стимул определенной частоты. Есть и свидетельства того, что настройка клеток находится под контролем обратной связи от улитки. Базилярная мембрана и ее волосковые клетки, таким образом, оказываются посложнее клавиш фортепьяно или пластинок ксилофона. Тем не менее, более столетия назад Гельмгольц сумел ухватить суть явления. Сложная электромеханическая настройка волосковых клеток с модуляцией через обратные связи накладывается на механизм максимального возмущения в специфической части базилярной мембраны, связанной с частотой поступающего звука.
Многие млекопитающие чувствительны к звукам намного больших частот, чем способно различить человеческое ухо. Эксперименты по выработке условных рефлексов при воздействии на вибриссы или ушную раковину показали, что многие мелкие млекопитающие чувствительны к звукам частотой до 100 кГц. Слышимый для человека писк мышей и землероек находится в низкочастотной части спектра слышимости этих животных. Большая часть социальной коммуникации между этими млекопитающими реализуется за пределами слышимости для человека. У собак частотная характеристика тоже получше, чем у человека — их верхний предел находится в районе 35 кГц. Это используют производители собачьих ультразвуковых свистков. А вот домашние кошки слышат почти, как мы1. Китообразные, с другой стороны, испускают и воспринимают звуки намного более высокочастотные — до 100 кГц. Такие звуки могут использоваться в социальной жизни вида, а у некоторых видов, напр, дельфина (Delphinus), и в эхолокации. Интенсивность и продолжительность эмиссии звука у них возрастает по мере приближения к препятствию или при поиске пищи2.
Date: 2015-10-18; view: 2051; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |