Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Научение как образование «следов» и как системогенез
Системно-селекционная концепция научения
Идея развития, наряду с идеей системности, относится к фундаментальным положениям теории функциональных систем. Обе они были воплощены в концепции системогенеза, которая сформулирована с привлечением обширного экспериментального материала, накопленного при исследовании формирования нервной системы и поведения. В этих исследованиях было обнаружено, что в процессе раннего онтогенеза избирательно и ускоренно созревают именно те элементы организма, имеющие самую разную локализацию, которые необходимы для достижения результатов систем, обеспечивающих выживание организма на самом раннем этапе индивидуального развития [12].
В настоящее время становится общепризнанным, что многие закономерности модификации функциональных и морфологических свойств нейронов, а также регуляции экспрессии генов, лежащие в основе научения у взрослых, сходны с теми, которые определяют процессы созревания, характеризующие ранние этапы онтогенеза [9,10]. Это дает авторам основание рассматривать научение как реактивацию процессов созревания, имеющих место в раннем онтогенезе.
В теории функциональных систем, наряду с признанием специфических характеристик ранних этапов индивидуального развития по сравнению с поздними [3,41], уже довольно давно [32,36] было обосновано представление о том, что системогенез имеет место не только в раннем онтогенезе, но и у взрослых. Формирование нового поведенческого акта в любом возрасте есть формирование новой системы - системогенез.
Позднее был сделан вывод о том, что принципиальным для понимания различий роли отдельных нейронов в обеспечении поведения является учет истории формирования поведения, т.е. истории последовательных системогенезов [3,5], а затем разработана системно-селекционная концепция научения ([147], см. также в [4,6,7,38]).
Системно-селекционной концепции созвучны современные идеи о «функциональной специализации», пришедшие на смену идеям «функциональной локализации», и о селективном (отбор из множества клеток мозга нейронов с определенными свойствами), а не инструктивном (изменение свойств, «инструктирование» клеток соответствующими сигналами) принципе, лежащем в основе формирования нейронных объединений на ранних и поздних стадиях онтогенеза [77]. Представление, сложившееся как в когнитивных науках, так и в биологии, о том, что индивид снабжен «многоцелевыми» врожденными элементарными «кирпичиками» - «примитивами», из которых в условиях инструктирующего влияния среды может быть сконструировано что угодно в процессе «ассоциативного обучения», подвергается разносторонней критике [126]. Так, например, Дж. Эдельманом доказательно обосновываются преимущества представления, согласно которому научение есть процесс селекции и прогрессивной специализации [77].
Принцип селекции по Эдельману может быть сведен к следующим положениям. В мозгу формируются группы нейронов, каждая из которых по-своему активируется при определенных изменениях внешней среды. Специфика группы обусловлена как генетическими, так и эпигенетическими модификациями, происшедшими независимо от упомянутых изменений. Селекция имеет место уже при созревании мозга в раннем онтогенезе, в процессе которого множество нейронов гибнет. Отобранные же клетки составляют первичный ассортимент. Вторичный ассортимент, формируется в результате селекции, происходящей при научении в процессе поведенческого взаимодействия со средой. Принятие положения о селекции как основе развития на всех его этапах устраняет дихотомию между созреванием и научением [134].
В рамках системно-селекционной концепции научения формирование новой системы рассматривается как формирование нового элемента индивидуального опыта в процессе научения. В основе формирования новых функциональных систем при научении лежит селекция нейронов из «резерва» (предположительно низко активных или «молчащих» клеток). Эти нейроны могут быть сопоставлены с первичным ассортиментом и обозначены как преспециализированные клетки.
Селекция нейронов зависит от их индивидуальных свойств, т.е. от особенностей их метаболических «потребностей». Отобранные клетки становятся специализированными относительно вновь формируемой системы. Эти нейроны могут быть сопоставлены с вторичным ассортиментом по Эдельману.
Специализация нейронов относительно вновь формируемых систем — системная специализация — постоянна. (Интересно, что сформированная «память» иммунных клеток также постоянна, они никогда не «забывают» [93].) Таким образом, новая система оказывается «добавкой» к ранее сформированным, «наслаиваясь» на них.
Положения о наличии в мозге животных разных видов большого числа «молчащих» клеток, об увеличении количества активных клеток при обучении, а также о том, что вновь сформированные специализации нейронов остаются постоянными (в эксперименте - в течение всего периода хронической регистрации: недели и даже месяцы), и что при научении происходит скорее вовлечение новых нейронов, чем переобучение старых, согласуются с данными, полученными в работах ряда лабораторий [59,63,99,152,156,160,162].
Актуализация памяти при реализации поведения
Как используется материал памяти, сформированный при научении, для реализации поведения? В экспериментах с регистрацией активности нейронов, специализированных относительно систем разного «возраста», обнаружено, что осуществление поведения обеспечивается не только посредством реализации новых систем, сформированных при обучении актам, которые составляют это поведение, но и посредством одновременной реализации множества более старых систем, сформированных на предыдущих этапах индивидуального развития. Последние могут вовлекаться в обеспечение многих поведений, т.е. относиться к системам, которые являются общими для разных актов. Замечу, что если один и тот же нейрон, принадлежащий к такой «общей» системе, вовлекается в разные акты, то характеристики его активации в этих актах различаются, так как в них он должен согласовывать свою активность с активностью разных наборов клеток [3].
Таким образом, системы, реализация которых обеспечивает достижение результата поведенческого акта, формируются на последовательных стадиях индивидуального развития. Системная структура поведения отражает историю его формирования. Иначе говоря, реализация поведения есть реализация истории формирования поведения, т.е. множества систем, каждая из которых фиксирует этап становления данного поведения. Отсюда логически следует и экспериментально подтверждается [6,17] следующее положение. Нейрональное обеспечение внешне одного и того же поведения различается, если история формирования этого поведения различна. А, следовательно, различается и структура памяти, актуализируемая (активируемая) при реализации этого поведения. Итак, поскольку история формирования поведения лежит в основе закономерностей его реализации, ясно, что «исторический нигилизм» отрицательно сказывается на понимании получаемого экспериментального материала.
Замечу, что «исторический нигилизм» может сказываться на понимании результатов исследования не только в тех случаях, когда анализируется нейронное обеспечение поведения. Содержание памяти всегда индивидуально. Даже в тех случаях, когда речь идет о видоспецифических актах. Это содержание определяет характер любого взаимодействия индивида со средой. Влияние особенностей индивидуального развития на активность мозга, свойства рецептивных полей нейронов, в том числе клеток первичных сенсорных областей коры, обнаруживается при решении самых разных исследовательских задач (см., например, [70,78,87,89,115,133,144]). Имея в виду методы экспериментирования аналитической физиологии, важно подчеркнуть, что эти особенности проявляется не только у бодрствующих индивидов. Они обнаруживает себя, например, в измененной чувствительности синапсов при исследовании срезов мозга [48] или при анализе «настройки» нейронов наркотизированных животных на те или иные параметры стимуляции [88,153].
Из сказанного следует, что описать механизмы поведения с системных позиций – значит выяснить, каковы «паттерны нейронных специализаций» [3,6] разных структур мозга, т.е. относительно каких конкретно систем специализированы нейроны этих структуры, как формируются системные специализации. Это значит также изучить динамику согласования активности нейронов, принадлежащих к одной и той же и к разным системам, т.е. динамику «внутрисистемных» и «межсистемных» отношений при реализации поведения. Я считаю, что рассмотрение упомянутых вопросов в качестве ключевых хорошо согласуется с тем набором проблем, которые выделял как центральные для системного подхода П.В. Симонов. Он подчеркивал, что «системный подход продуктивен только в том случае, если мы более или менее определенно указываем на конкретные элементы, из которых состоит данная система, на функциональное значение каждого из этих элементов и на правила взаимодействия элементов друг с другом при функционировании системы в целом» [30, с.95].
Неонейрогенез
К настоящему времени получены убедительные доказательства наличия неонейрогенеза у взрослых млекопитающих, в том числе у людей [82,129]. Получены также данные о том, что, хотя синаптическая активность появляется у этих новых нейронов довольно быстро после завершения миграции, но спайковая активность у них отсутствует; из всех свойств нейрона она оказывается последней по времени появления [61]. Следовательно, в динамике вовлечения вновь появившихся клеток в совместную деятельность наблюдаются переход от «молчания» к импульсацции, как это, возможно, происходит при вовлечении «резервных» клеток в процесс специализации. Упомянутые данные, наряду с результатами, полученными G.Kempermann et al. [103], показывающими, что число выживших нейронов, вновь появившихся в процессе нейрогенеза у взрослых животных, увеличивается при содержании последних в обогащенной среде (принцип «используй или потеряешь» [129]), а также данные о том, что искусственное угнетение нейрогенеза нарушает формирование памяти [146], позволяют высказать следующее предположение: неонейрогенез может вносить вклад в процессы системогенеза (рис. 2). Наряду с рекрутированием клеток «резерва» и вновь появившиеся нейроны специализируются относительно новых систем. Как мне представляется, дополнительным сильным аргументом в пользу высказанного предположения являются данные, полученные в лаборатории F.N. Nottebohm [124]. Было обнаружено, что у канареек нейроны вентрального гиперстриатума, специализированные относительно новой песни (их метили пероксидазой хрена), имеют и тимидиновые метки, относясь к вновь появившимся нейронам.
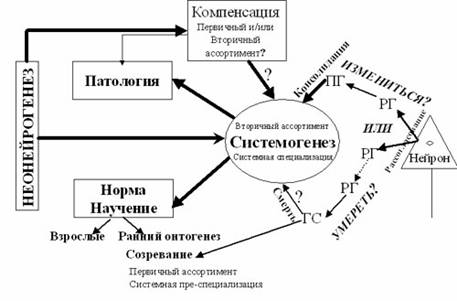
Рис. 2. Умереть или измениться? Теоретическая схема.
РГ – «ранние» гены, ПГ – «поздние» гены, ГС – гены «смерти». Подробности – см. текст.
Неонейрогенез может иметь значение и для замены нейронов первичного и/или вторичного ассортиментов, гибнущих как в условиях нормы, так и при патологии (см. ниже). Что касается патологии, в литературе уже имеются данные об интенсификации неонейрогенеза при локальных поражениях мозга взрослых индивидов и о миграции вновь появившихся нейронов в область повреждения [163]. Знаки вопроса на рис. 2 (внутри фрагмента Компенсация и у стрелки, идущей от него) означают, что в настоящее время трудно сказать, какой именно ассортимент (или оба) подлежит восполнению, и каким образом мог бы меняться состав ранее специализировавшихся нейронов в том случае, если компенсация вторичного ассортимента имеет место.
С пополнением набора преспециализированных клеток (первичного ассортимента) ситуация кажется более ясной. Если он имеет место, то, по-видимому, «встраивание» новых нейронов должно происходить по общим для всех преспециализированных клеток «резерва» правилам – в процессе системогенеза. Если таковой не происходит в течение определенного времени, вновь появившиеся нейроны гибнут. Я считаю, что в пользу этого говорят многочисленные данные, подобные приведенным выше данным G.Kempermann.
Говоря о пополнении набора преспециализированных клеток (первичного ассортимента) за счет нейрогенеза в норме, можно отметить, что в настоящее время имеются веские основания думать, что научение ведет не только к выживанию уже появившихся нейронов, но и интенсификации пролиферации. Предполагается, что эта интенсификация имеет значение для будущего научения [129].
Что касается компенсации специализированных клеток (вторичного ассортимента), трудно допустить, чтобы и она могла происходить вне системных процессов, требующих активации клеток, принадлежащих к системе, и проб, направленных на достижение ее результата. Возможно такие пробные согласования новых и «старых» клеток могут развертываться во время реконсолидационных процессов (см. ниже).
Консолидация памяти: от формирования и упрочения «следов» посредством повышения эффективности синапсов к системогенезу
Проблема формирования и консолидации памяти решается с применением самых современных методов и базируется на современных концептуальных схемах. Несмотря на это, в основе большинства этих схем и исследований – идея Декарта, сформулированная им более трехсот лет назад: «следы» существуют потому, что поры в мозгу, через которые дух проходил раньше, становятся все более проходимыми, когда дух проходит через них снова. И дух легче проходит именно через эти поры. В соответствии с этой идеей важно выяснить, каковы механизмы увеличения проходимости пор, в каких мозговых структурах больше подобных пор, все ли поры обладают свойством изменчивости, сколь долго сохраняется повышенная проходимость пор и т.д., и т.п. Соответственно, при всем разнообразии подходов к пониманию консолидации, в качестве наиболее базовой ее закономерности рассматривается долгосрочное усиление синаптического проведения в дуге (ах) рефлекса, сетях и пр.
Иначе выглядит процесс формирования и упрочения памяти с системных позиций. С этих позиций, как уже было отмечено, нейрон не является проводником возбуждения. Поэтому проблема увеличения эффективности проведения не возникает. Формирование нового поведения рассматривается не как проторение путей и образование «следов», осуществляемое за счет увеличения синаптической эффективности. Не как связывание структур, последовательное активирование которых обеспечивает реализацию вновь сформированного поведения. Оно представляет собой формирование новой системы совместно активирующихся клеток организма, в том числе и нейронов, расположенных в самых разных структурах мозга, вовсе не обязательно связанных прямыми влияниями.
Переход от первой ко второй, системной позиции осуществляется в настоящее время уже не только на основе теоретических соображений, но, дополнительно, исходя из результатов экспериментов. В недавней обзорно-теоретической статье G. Horn отмечает, что результаты кросскорреляционного анализа активности нейронов IMHV не подтверждают представления о том, что при научении происходит усиление связей между нейронами, как это должно было бы быть при образовании «хеббовских ансамблей». «Скорее, - заключает автор – нейроны формируют набор параллельно [организованных], в значительной степени прямо не связанных элементов, что обеспечивает большую эффективность в сохранении [памяти], чем система непосредственно связанных элементов» [95, p.121].
Полагаю, что подобная позиция приходит в серьезное противоречие с базовыми положениями традиционного подхода. И хотя обоснованные призывы «пересмотреть имеющиеся модели [консолидации] и не быть при этом догматичными в отношении взглядов, не совпадающих с нашими предвзятыми представлениями о работе мозга» [116, p.467] имеют место, но все же внутри традиционного подхода позиция G. Horn пока может рассматриваться как оригинальная. В том же журнале, в котором появилась цитированная выше статья G. Horn, тремя месяцами раньше, была опубликована дискуссия, участник которой J. Lisman писал: «Как считал Хебб, ансамбли формируются, когда синаптические связи между активными клетками избирательно усиливаются. Я не слышал ни о каких обоснованных альтернативах, объясняющих сохранение ассоциативной памяти» [110, p.929].
Какие же закономерности можно выделить в процессах формирования и сохранения памяти, если посмотреть на них с системной точки зрения? И если не рассматривать в качестве основы таких процессов механизм увеличения синаптической проводимости в цепях связанных нейронов, как быть с огромным материалом, полученным при изучении таких механизмов? К такому материалу относятся, в частности, многочисленные работы, демонстрирующие активацию генетического аппарата клетки, изменение возбудимости и морфологические перестройки нейронов при научении. Для ответа на первый вопрос необходимо сначала сказать несколько слов о реконсолидации.
Реконсолидация при реактивации памяти и при научении
Еще F. Bartlett [54] предлагал полностью отбросить взгляды, в соответствии с которыми «воспроизведение из памяти» рассматривается как «повторное возбуждение неизменных «следов». В настоящее время обнаружены молекулярно-биологические закономерности реконсолидации памяти, лежащие в основе ее модификации после повторной актуализации (см., например, [137]). При формировании нового материала памяти необходим процесс синтеза белков, лежащий в основе процессов ее консолидации. Активация памяти, как и ее формирование, требует синтеза белка для реконсолидационных процессов. В связи с этим в последнее время предлагается связывать протеин-зависимые консолидационные процессы не с «новой», а шире - с «активной» памятью [117].
Представление о реконсолидационных модификациях не вступают в противоречие с приведенным выше положением о постоянстве системной специализации нейронов, которая формируется при научении. Анализ накопленного экспериментального материала позволяет K.Nader заключить, что, конечно, данные о реконсолидации «противоречит взгляду на память как лабильную только один раз». Однако он специально подчеркивает, что «реактивация консолидированной памяти не отменяет модификаций, обусловивших формирование долговременной памяти (вне зависимости от того, какие они - структурные или химические)» [116, р.467].
Реконсолидационные процессы представляют для нас особый интерес в связи с тем системным представлением о формировании памяти, которое было изложено выше. С точки зрения системно-селекционной концепции, оно представляет собой специализацию новой группы нейронов относительно вновь формируемой системы и «добавление» последней к ранее сформированным системам. Это добавление, видимо, требует взаимного согласования нового элемента и ранее сформированных и приводит к модификации последних.
Поведенческие данные, полученные в лаборатории И.П.Павлова [24], позволили ему прийти к заключению о том, что прибавление новых условных рефлексов сейчас же отзывается на состоянии прежних. В последнее время на основании данных, полученных в экспериментах с определением системной специализации нейронов при последовательном формировании разных поведенческих актов, также был сделан вывод об изменении ранее сформированной системы поведенческого акта после обучения следующему акту. В экспериментах с регистрацией нейронной активности у животных, первоначально обученных инструментальному пищедобывательному поведению, а затем в той же экспериментальной клетке алкогольдобывательному поведению, было обнаружено, что нейроны, специализированные относительно систем первого поведения, претерпевают при формировании второго модификацию и начинают вовлекаться также и в обеспечение алкогольдобывательного поведения вместе с нейронами, вновь специализировавшимися относительно этого поведения. Эта реконсолидационная модификация, претерпеваемая предсуществующей, «старой» системой при появлении связанной с ней новой системы, была названа «аккомодационной» реконсолидацией [47].
Естественно, такая позиция предполагает постановку вопроса о сходстве и различиях между реактивационной и аккомодационной реконсолидациями. Экспериментальная разработка этой проблемы имела бы большое значение для формулировки более дифференцированных представлений о формировании и функционировании памяти.
Консолидация с системной точки зрения
Сказанное выше, как мне представляется, означает, что при анализе формирования памяти следует учитывать не только появление ее новых элементов, но и модификацию ранее сформированных. При учете этого фактора можно предложить системное описание процесса консолидации. Консолидация с этой точки зрения включает две группы неразрывно связанных процессов.
1) Процессы системной специализации: морфологическая и функциональная модификация нейронов, связанная с их вовлечением в обеспечение вновь формируемой системы.
2) Процессы аккомодационной реконсолидации, обусловленные включением этой системы в существующую структуру памяти индивида: морфологическая и функциональная модификация нейронов, принадлежащих к ранее сформированным системам.
Обнаруживаемые во множестве нейрофизиологических, морфологических, молекулярно-биологических и других исследований модификации нейронов, сопутствующие научению, могут быть связаны как с первой, так и со второй группой процессов. Так, например, появление активаций на условный сигнал у тех нейронов, которые до сочетания условного и безусловного сигналов отвечали только на безусловный, связаны скорее всего именно с аккомодационной реконсолидацией, а не с системной специализацией (см. также [3]). То же можно сказать относительно модификаций в первичной моторной коре, в которой после обучения разным поведенческим актам выявляется преобладание нейронов, специализированных относительно «старых» систем, сформированных на ранних этапах индивидуального развития [1,6]. Именно с реорганизацией этих нейронов, видимо, связано увеличение уровня экспрессии «ранних» генов (подробнее см. ниже) при обучении крыс инструментальному нажатию на педаль лапой, обнаруживаемое в зоне проекции этой лапы в первичной моторной коре [62].
Дифференцированный подход к модификациям первой и второй группы явится существенным шагом вперед на пути к пониманию закономерностей формирования памяти.
В начале формирования памяти: экспрессия «ранних» генов как показатель рассогласования
Консолидация, с какой бы точки зрения ее ни рассматривать, включает морфологические изменения нейронов (см., однако, [50]), такие, например, как изменение размера синапсов, изменение их числа [53]. Начальным звеном каскада молекулярно-биологических процессов, обусловливающих морфологические модификации нейронов как в процессе морфогенеза (ранний онтогенез), так и при консолидации памяти, формируемой у взрослых индивидов, является экспрессия «ранних» генов. Активация «ранних» генов довольно кратковременный процесс (занимает около 2 часов), сменяемый второй волной экспрессии - «поздних» генов; в составе второй волны активируются морфорегуляторные молекулы, имеющие непосредственное отношение к морфологическим модификациям нейрона [9,10].
В настоящее время связь экспрессии «ранних» генов с процессами научения, которая отмечалась уже давно (см. в [9,10]), становится все более очевидной [95]. В системно-селекционной концепции научение связывается с формированием специализаций нейронов относительно формируемой системы. Логично было бы тогда ожидать, что экспрессия «ранних» генов и формирование специализаций связаны между собой. В исследованиях, проведенных нашей лабораторией совместно с отделом системогенеза НИИ нормальной физиологии им. П.К. Анохина [29], были получены данные, указывающие на то, что экспрессия «ранних» генов лежит в основе формирования специализаций нейронов в отношении вновь формируемых инструментальных поведенческих актов. В тех мозговых структурах, в которых после обучения обнаруживалось достоверно больше вновь специализированных нейронов, была соответственно достоверно более выражена и экспрессия (рис. 3; данные экспериментов О.Е. Сварник и др. [29]).
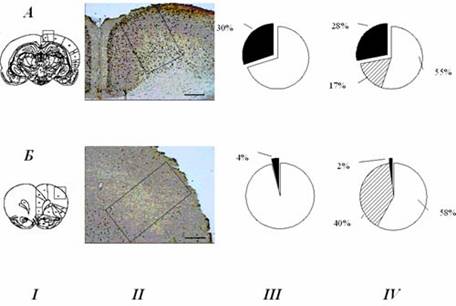
Рис. 3. Сопоставление относительного числа Fos-положительных клеток (III) с паттернами поведенческих специализаций нейронов (IV) в цингулятной (А) и антеролатеральной (Б) коре.
I - диаграммы фронтальных срезов, обозначающие исследуемые области. II - микрофотографии фронтальных срезов, показывающие Fos-окрашенные ядра нейронов крысы, сформировавшей новое поведение; калибровка: 500 мкм. III - темный сектор – процент нейронов, экспрессирующих c-Fos; светлый сектор – не экспрессирующих c-Fos. IV - темный сектор – процент нейронов, принадлежащих к «новым» системам, сформированным при обучении крыс инструментальному акту нажатия на педаль в пищедобывательном поведении; сектор со штриховкой – процент нейронов, принадлежащих к «старым» системам, сформированным на этапах индивидуального развития, предшествующих обучению инструментальному пищедобывательному поведению; светлый сектор – нейроны, не вовлекающиеся в обеспечение данного поведения. Видно, что в цингулятной коре значительно выше, чем в антеролатеральной, как число клеток, экспрессирующих c-Fos, так и число нейронов, специализированных относительно вновь сформированной системы инструментального поведенческого акта.
Активация «ранних» генов у взрослого индивида имеет место не только при научении, но и при голоде, стрессе, поражениях нервной системы или ишемии мозга [74,123,150]. Показано, что изменение микросреды нейронов обусловливает появление активности у ранее молчавших клеток [40] и экспрессию ранних генов [57,150]. Поэтому, имея в виду сказанное выше о детерминации активности нейрона рассогласованием между его «потребностями» и притоком метаболитов, а также аргументированную возможность рассмотреть экспрессию ранних генов как специфическое проявление активности клетки [65], возникающей в ситуации новизны [11], логично предполагать, что общим для всех перечисленных выше ситуаций, включая научение, является рассогласование. Оно возникает вследствие того, что ранее сформированные способы согласования метаболических «потребностей» нейронов оказываются неэффективными в условиях стойкого изменения микросреды нейронов.
Рассогласование является, в известном смысле, межпарадигмальной концепцией, важнейшим компонентом не только теории функциональных систем, но и других теорий. Так, П.В.Симонов отмечал «генетическое родство» биологической и информационной теории эмоций и подчеркивал, что обе они восходят к одному «идейному источнику – к концепции Павлова о несовпадении (рассогласовании) ранее сложившегося внутреннего и изменившегося внешнего динамического стереотипа» [30, c.60].
Поиск новых путей устранения рассогласования включает как модификации на уровне поведенческих адаптаций, так и молекулярно-генетические и морфологические перестройки. Очевидно, что подобные модификации имеют место как в норме, так и в патологии.
Хотя в «обыденной науке» (folk science; основанное на здравом смысле, не требующее точных определений понимание) норма и патология мыслятся как принципиально различные состояния, причем патология рассматривается как слом, разрушение нормальных отлаженных механизмов, но в собственно науке уже давно обосновано представление о том, что процессы, называемые патологическими, не являются дезорганизацией или хаосом, а лишь своеобразным, выработанным в процессе эволюции приспособлением к условиям существования; нет ни одного патологического процесса, который не имел бы своего прототипа в норме [14,19,90].
О формировании новой системы, направленной на достижение конкретного результата, как о важнейшем звене механизмов научения уже шла речь выше. Адаптационные изменения организации внутренней среды и соотношения индивида со средой внешней, возникающие в условиях патологии, могут быть также рассмотрены, как системный процесс, захватывающий весь организм и направленный на обеспечение возможности достижения положительных результатов [13,90]. Многочисленными исследованиями динамики активности мозга после локальных его повреждений показано, что в интактных структурах мозга развиваются процессы реорганизации, обусловливающие восстановление поведения, и что эти процессы сопоставимы с пластическими перестройками, имеющими место при научении в норме [46,68,158].
Так, в настоящее время накапливается все больше данных, позволяющих полагать, что между нейронными механизмами, лежащими в основе формирования долговременной памяти при обучении и в основе «долгосрочных адаптаций», возникающих при хроническом воздействии аддиктивных веществ, имеется существенное сходство [120,135]. Результаты наших исследований позволили предположить, что определенный вид «долгосрочных адаптаций», имеющих место при хроническом употреблении алкоголя, не просто сходен, но идентичен модификациям, лежащим в основе формирования нового опыта. К ним относятся перестройки нейронов, связанные с формированием новых специализаций относительно алкогольдобывательного поведения, а также с процессами аккомодационной реконсолидации преморбидных (сформированных до возникновения потребности в алкоголе) специализаций [47].
В связи с только что отмеченным сходством, а также при учете сходства молекулярно-биологических механизмов, лежащих в основе созревания и научения, не удивительно, что реювенилизация - активация у взрослого процессов, характерных для созревания мозга в раннем онтогенезе, - имеет место не только при научении в норме, но и при восстановлении после поражений мозга [69]. Для целей дальнейшего обсуждения важно подчеркнуть, что к механизмам, реактивируемым в условиях патологии у взрослого, относят и апоптотическую гибель клеток: физиологический апоптоз в развивающемся мозге и патологический апоптоз во взрослом мозге имеют сходные молекулярные механизмы [164].
Нейрон, как отмечалось, может обеспечить «потребности» своего метаболизма, объединяясь с другими элементами организма в функциональную систему, извлекаемую из памяти. Достижение результата системы устраняет рассогласование между «потребностями» и микросредой нейронов. Подобная динамика характеризует ситуацию дефинитивного поведения. Ситуация научения в норме и восстановления в патологии (например, после инсультов, травматических и т.п. поражений мозга) специфична тем, что проблему согласования «потребностей» нельзя решить с использованием имеющихся у индивида способов согласования (т.е. в рамках имеющегося у индивида опыта). Рассогласование в данной ситуации отличается от того, которое имеет место в дефинитивном поведении: оно устраняется поиском и фиксацией в памяти новых вариантов объединения элементов, развертыванием процессов системогенеза (рис. 2).
Таким образом, имея в виду научение и слегка перефразируя утверждение Сократа, можно резюмировать содержание данного раздела следующим образом: рассогласование есть начало всей мудрости.
От рассогласования через согласование к консолидации
В случае если процессы системогенеза протекают успешно, как уже отмечалось, формируются новые системы, устанавливаются и консолидируются новые межнейронные отношения, обеспечиваемые, в том числе, морфологическими перестройками (в основе которых - активация генетического аппарата) и обеспечивающие удовлетворение метаболических «потребностей» нейронов, а, следовательно, и их выживание.
Как выглядит поиск путей согласования метаболизмов клеток в процессе обучения при анализе «внешнего» поведения или электрической (суммарной и импульсной) активности мозга? На уровне поведения, если это не особый случай обучения с одной реализации, могут быть отмечены ориентировочно-исследовательское поведение, пробы, не приводящие к достижению полезного результата, а затем завершающиеся его достижением. Однако даже достижения результата не означают, что процессы научения завершены. Анализ актограмм, числа ошибок и пр. демонстрирует динамику. Через некоторое время, разное для разных видов поведения, показатели поведения стабилизируется. Следует заметить, что впечатление стабилизации пропадает, если рассматривать поведение под большим «увеличением». Каждый новый акт – не просто повторное извлечение из памяти, а созидание. Поведенческие акты обнаруживают направленную динамику (совершенствуются) в течение тысяч реализаций [92]. Изменчивость поведения при повторных реализациях может быть исследована и на нейронном уровне [8].
Что касается суммарной активности мозга, регистрируемой у человека, показано, что она претерпевает изменения не только в процессе обучения навыку, но и через часы (и дни) после того, как по поведенческому критерию испытуемый уже обучился [102,142]. Иначе говоря, скажем, через 30 минут и через 5 часов с момента начала реализации только что выученного поведения, активность мозга существенно различается, несмотря на то, что результативность поведения в процессе реализации существенно не меняется. В экспериментах на животных также было показано, что как характеристики активаций нейронов, так и число активирующихся клеток меняются от первых реализаций поведения в течение последующих часов и дней [39,64,81,86,95,96,99,111,119,131,140]. Число клеток изменяется нелинейно. Направленность изменения зависит, по-видимому, от характера формируемого поведения, структуры, в которой регистрируется активность нейронов, и этапа, на котором регистрация производится.
Данные, полученные О.Е. Сварник [28], показывают, что число клеток мозга, в которых наблюдается экспрессия ранних генов, во много раз превышает число нейронов данной области, у которых обнаруживается специализация относительно системы формируемого поведения. Мы предполагаем, что часть из множества генетически активированных клеток – это нейроны, специализированные относительно систем ранее сформированных актов, и экспрессия в них отражает начало процесса аккомодационной реконсолидации. Большинство же этих клеток являются преспециализированными, и их генетическая активация является предпосылкой перехода клеток в состояние готовности для селекции в процессе проб. Именно в процессе проб из активированных (как генетически, так, вероятно, и импульсно) клеток отбираются те, которые перейдут далее в разряд специализированных относительно формируемой системы.
Отражением протекания описанной селекции и изменений, происходящих в нейронном обеспечении вновь сформированного поведения, является обнаруженное в наших пилотажных экспериментах (Е.А. Кузина и др.) изменение вариативности связи нейронов цингулятной коры крыс с новым поведением. Отмечена тенденция к увеличению числа нейронов, у которых активация появлялась в 100% случаев (т.е. при каждой реализации специфического для данной клетки акта) при сравнении совокупностей нейронов, активирующихся в инструментальном пищедобывательном поведении в течение первых дней и через 7-15 дней после завершения обучения. По-видимому, это изменение активности, уменьшение ее вариативности по мере консолидации памяти, может быть связано с завершением селекции и стабилизацией состава нейронов, вовлекающихся во вновь сформированное поведение. Полученные результаты мы рассматриваем как предварительные, но они согласуются с результатами экспериментов других авторов. Ranganath C. и Rainer G. [131] описывают данные, полученные при регистрации активности нейронов префронтальной коры у обезьян, которым предъявляли новые и знакомые объекты. Хотя предъявление нового объекта вызывает активации у большего числа нейронов, чем предъявление знакомого объекта, но в последнем случае активации существенно более четкие.
Однако данные литературы не позволяют свести объяснение изменения состава активирующихся нейронов только к описанной селекции. Например, не исключено, что некоторые нейроны могут активироваться на начальной стадии обучения, затем прекратить активность, а затем вновь начать разряжаться [95]. В то же время показано, что часть клеток начинают активироваться на сравнительно поздних этапах обучения, лишь в период стабилизации поведения (см., например, [161]), в то время как другие клетки, и это особенно важно для настоящего анализа, активируются лишь на начальных стадиях обучения, а когда поведение стабилизируется, их активность прекращается и больше не возникает [161,144]. Одно из возможных и, как мне кажется, вполне правдоподобное объяснение этого феномена сводится к следующему.
Более 30 лет назад J. B. Ranck [130] описал в гиппокампе группу клеток, которые активировались лишь в ситуации, когда в среде, к которой животное было адаптировано, происходили изменения: появление нового объекта или исчезновение привычного. В других областях мозга также были описаны нейроны, активации которых оказывались специфически связанными с ситуацией новизны. К структурам, особенно чувствительными к упомянутой ситуации, кроме уже названного гиппокампа, относят зрительную, энторинальную, нижнюю височную, префронтальную, цингулятную, моторную и премоторную области коры, миндалину и др. [131].
Сопоставляя эти наблюдения с только что упомянутыми данными о существовании нейронов, активирующихся лишь в начале обучения (ситуация новизны), можно предположить, что первые пробные акты осуществляются за счет коактивации переменных составов преспециализированных нейронов и нейронов, которые пока условно назовем нейронами «новизны». Именно их коактивация обеспечивает как совершение проб, так и достижение при обучении первых положительных результатов. После стабилизации поведения нейроны «новизны» прекращают активность. Это означает серьезную реорганизацию структуры актуализируемой памяти и, возможно, инициирует следующую, дополнительную волну селекции.
В рамках традиционного подхода можно было бы говорить о том, что роль этих нейронов – обеспечить внимание, повысить уровень бодрствования и пр. и тем самым воздействовать на обучающиеся нейронные сети. Однако с позиций последовательного системного подхода такие и им подобные объяснения не могут быть приняты.
Как было отмечено выше, поведение осуществляется за счет реализации множества систем. При этом активация нейрона – показатель того, что в данном поведении актуализирована система, к которой он принадлежит. С этих позиций «новизна» - это не описание специализации нейронов, а лишь феноменологическое описание свойств их активности (ничем, правда, не худшее, чем огромное множество других: сенсорные нейроны, моторные нейроны, нейроны места, нейроны сознания и пр.). Системное же описание требует указания систем, сформированных для достижения определенного результата, к которым данные нейроны принадлежат.
П.В. Симонов считал окончательно доказанным существование у животных исследовательского поведения, которое побуждается «самостоятельной потребностью» в обеспечении контакта с объектами, «прагматическое значение» которых не выяснено [31]. Для достижения каких результатов формируются системы упомянутого поведения?
Память может быть разделена на домены. В.Б. Швырков [37] подчеркивал, что то, элементы какого именно домена памяти будут актуализированы индивидом в поведении, определяется как мотивацией, так и тем, в какой среде он находится. Наиболее глобальное разделение доменов подразумевает противопоставление памяти, сформированной для осуществления поведений «приближения» и «избегания» («approach» и «withdrawal» [138]; см. подробнее в [44,56]).
Когда индивид попадает в незнакомую ситуацию, ситуацию с элементами неопределенности, первоочередной задачей является достижение следующего результата: отнесение ситуации к тем, которые позволяют совершать одно из поведений группы «приближения», или предполагают поведение «избегания». Это означает возможность или необходимость актуализации элементов памяти того или другого домена. Достижение упомянутого результата позволяет перейти к осуществлению поведения, направленного на достижение следующего результата в поведенческом континууме. Каков будет этот следующий результат, определяется и тем, какова мотивация индивида, и тем, каков результат исследовательского поведения, т.е. тем, элементы какого домена могут быть актуализированы. Можно полагать, что необходимость решения подобных задач существует и при устранении неопределенности, осуществляемой как выбор «внутри» упомянутых больших доменов, т.е. выбор между субдоменами. Нельзя исключить, что авторы классифицируют как «исследовательское» также и то поведение, реализуемое индивидом, которое в сходной, с точки зрения индивида, ситуации (first person perspective), приводило ранее к достижению полезного результата. Каков был этот результат, может быть неизвестно наблюдателю. Вероятно, по крайней мере, часть из нейронов, активирующихся лишь на начальных этапах процесса формирования памяти, принадлежат к системам исследовательского поведения, актуализируемым в процессе обучения вместе с создаваемыми системами нового поведения.
По-видимому, динамику описанных в настоящем разделе процессов возникновения рассогласования и его устранения за счет установления новых межклеточных отношений и нового соотношения организма и среды характеризует и феномен долговременной потенциации (ДП; LTP). Однако для того, чтобы обосновать возможную системную трактовку этого феномена, необходимо прежде остановиться на описании ситуации затянутого рассогласования.
Элиминация нейронов как «альтруистичный суицид»
Если рассогласование между «потребностями» нейронов и их микросредой не устранено, нейроны гиперактивны, экспрессия «ранних» генов затягивается: одна волна экспрессии сменяет другую. В этих случаях в нейронах могут экспрессироваться так называемые гены «смерти», активация которых ведет к гибели нервных клеток (рис. 2). Существующие экспериментальные данные рассматриваются как серьезный аргумент в пользу утверждения о связи между затянутой экспрессией «ранних» генов и программируемой смертью нейронов. Напротив, сравнительно краткосрочная экспрессия возникает у нейронов, которые выживают [141].
Итак, при наличии у организма опыта удовлетворения данных «потребностей» в данной ситуации, избирательно активируется память, имеющая отношение к удовлетворению данного набора «потребностей», и возникает импульсная активность нейронов, специализированных относительно актуализируемых элементов памяти - систем. Эта активность и есть нейронные основы реализации поведения. Достижение результата поведения на уровне соотношения целостного индивида и среды прекращает поведение, направленное на достижение этого результата, а на уровне отдельного нейрона оно выступает как устранение рассогласования между «потребностями» нейрона и микросредой.
В случае же отсутствия подобного опыта, когда повторные импульсации коактивированных нейронов не приводят к достижению результата, возникает экспрессия «ранних» генов. Эта экспрессия может быть рассмотрена как предпосылка для активации других транскрипционных компонентов – основы принимаемого клеткой «решения жить или умирать» [108, p.2736].
Следовательно, в случае возникновения рассогласования между «потребностями» нейрона и его микросредой и при невозможности устранить рассогласование в рамках имеющегося опыта, как в норме (в раннем онтогенезе и у взрослого), так и в патологии у него имеется, образно говоря, следующая альтернатива: измениться, вовлекаясь в системогенез (формирование новой системы, затем консолидируемой) или умереть (рис. 2). Вовлечение может носить характер системной специализации или аккомодационной реконсолидации (см. выше).
Излагаемые здесь соображения, в основе которых – представление об активности нейрона как детерминированной «потребностями» в метаболитах, согласуются с данным о том, что программируемая клеточная смерть запускается в условиях отсутствия соответствующих «факторов выживания» [132,164]. В то же время упомянутые представления не предполагают, что какое-то вещество всегда и при любых условиях производит одинаковый эффект. Напротив, ясно, что этот эффект должен зависеть от текущих «потребностей» клетки. Одно и то же вещество может как удовлетворить их в одном состоянии, так и привести к рассогласованию, если «потребности» изменились. Неудивительно поэтому, что трофические факторы, рассматриваемые как сигнал выживания, в определенном состоянии клетки могут превращаться в сигнал рассогласования и запускать программу клеточной смерти [164].
В рамках излагаемых представлений множественные повторные волны экспрессии «ранних» генов на начальных стадиях онтогенеза [118] возможно связать как с интенсивным морфогенезом и формированием все новых поведенческих актов (у многих животных за первые недели постнатального онтогенеза формируется больше половины актов всего поведенческого репертуара; см. в. [3]), так и с гибелью в этот период множества нервных клеток [118] (см. рис. 2).
Второй вариант развития событий, составляющих альтернативу «измениться или умереть», - смерть клеток - часто имеет место при созревании, а также в условиях патологии, при кардинальных изменениях микросреды клеток, в условиях, в которых оказывается невозможно использовать имеющийся у индивида опыт согласования метаболизмов клеток организма. Но не только в патологии и при созревании. Имеются данные, свидетельствующие в пользу апоптоза в мозгу здоровых взрослых индивидов, а также в пользу большого значения апоптоза нейронов нервной системы взрослого для функционирования целого организма [66,75,97,118,109,148].
Эти данные при учете представления о том, что системогенетические закономерности являются общим принципом реализации процессов созревания, научения на любом этапе онтогенеза, адаптации и восстановления в патологии, позволяют предположить, что упомянутая альтернатива существует и в норме. И что элиминация нейронов как один из исходов нейроселекции в раннем онтогенезе, значение которой для формирования поведенческого репертуара не вызывает сомнений, вносит вклад и в системогенез у взрослого (рис. 2; Знак вопроса на схеме у слова «смерть» обозначает гипотетичность представления о смерти нейронов как факторе системогенеза.).
Следовательно, формулируемая позиция сводится не к альтернативе «системогенез или смерть», а, коротко говоря, к двум взаимосвязанным путям обеспечения системогенеза: модификация нейрона или его гибель. Блокирование любого из них нарушает системогенетические процессы. Таким образом, здесь подчеркивается именно позитивный, в общеорганизменном плане, аспект гибели нейронов.
Фатальный для отдельных клеток исход - гибель – можно представить себе в качестве неизбежной платы за возможность осуществления успешного системогенеза на протяжении всего индивидуального развития. Неизбежной по крайней мере, в тех случаях, когда метаболические «потребности» каких-либо клеток вступают в неустранимое противоречие с новыми способами согласования «потребностей» клеток индивида. Формирование этих способов диктуется необходимостью соответствовать изменившимся условиям внешней и/или внутренней среды и выражается в образовании новых систем и изменении межсистемных отношений.
Можно предположить также, что особенно выраженное нарастание частоты смерти нейронов обнаружится при таком обучении, когда индивид долго не способен решить сравнительно сложную проблему, в частности, когда индивид оказывается в «неизбегательной» ситуации, что в субъективном плане может выражаться в депрессивном состоянии.
Выше представление об активном нейроне было противопоставлено представлению о нейроне реактивном. Здесь я хотел бы подчеркнуть, что принцип активности распространяется на весь период и на все аспекты существования нейрона, включая и процессы, связанные с реализацией альтернативы: измениться или умереть. Данная позиция находится в соответствии с точкой зрения о том, что каждый из этапов элиминации клетки является активным [132] и что, по существу, элиминация является суицидом [109,164].
Можно добавить, что этот суицид альтруистичен. В том смысле, что клетка включает программу самоэлиминации для того, чтобы таким образом устранить метаболическое противоречие, которое неустранимо другим путем, и обеспечить успешную адаптацию индивида к изменившимся условиям. А значит, обеспечить выживание других клеток, принадлежащих к тому же клону.
Ранее другими авторами уже приводились аргументы в пользу существования «альтруистичного клеточного суицида» в нервной системе: при инфицировании нейротропным вирусом. Альтруистичный суицид рассматривается как стратегия мультиклеточного организма, ограничивающего размножение вируса путем самоуничтожения инфицированных клеток [49].
У альтруизма клеток многоклеточного организма имеются эволюционные предпосылки. Описана альтруистическая гибель у одноклеточных (амебы Dictyostelium discoideum), которые приносят себя в жертву другим клеткам своего клона (чужого – менее охотно), обеспечивая существование временно формирующегося многоклеточного образования за счет формирования нежизнеспособного стержня, вокруг которого организуется колония. Остальные (около 80 %) клетки превращаются в жизнеспособные споры, составляющие это образование [151].
Долговременная потенциация: полезный артефакт?
Начну этот раздел с формулировки, отражающей мое понимание феномена ДП, а затем приведу аргументы в пользу этого понимания, рассмотрев ДП с позиций системного подхода: ДП – артефакт, который по не предусмотренным его исследователями причинам может иметь отношение к механизмам научения и памяти.
Долговременная потенциация синаптической эффективности (LTP) в результате тетанического раздражения проводящих путей в гиппокампе считается претендентом на роль физиологического механизма долговременной памяти и рассматривается в качестве экспериментальной модели пластичности, зависимой от активности. В ряде исследований, (см., например, [17,91,104,107,114]), показано, что ДП возникает не только в гиппокампе, но и в корковых структурах, не только при исследовании препаратов, но и у бодрствующих свободно подвижных животных.
В той области знания, которая является предметом анализа в настоящем сообщении, исследования ДП рассматриваются в течение ряда лет как наиболее важные и приоритетные. И это не только потому, что данный феномен хорошо укладывается в традиционную схему, описывающую формирование памяти как повышение синаптической эффективности в рефлекторных дугах. J. W. Lichtman и J.R. Sanes [110] отмечают существование также и факторов, которые они называют «социологическими». «Главной наградой для многих ученых является публикация в журналах с наиболее высоким [импакт фактором], – пишут авторы - [и ДП] как раз та область исследований, которая используется этими журналами, чтобы протрубить о новых данных». Однако далее они отмечают, что «пока будет существовать именно такая награда, до тех пор она будет порождать скорее поддержание дискуссий, но не решение проблемы», инициируя бесконечное обсуждение того, что «[природа] ДП – пресинаптическая; [природа] ДП – постсинаптическая; нет, пресинаптическая; нет, постсинаптическая; эта киназа вовлечена; та киназа вовлечена...» [110, p.928]. К упомянутым факторам можно добавить еще и следующий: «поддержание интереса к ДП, несмотря на раздражающую критику со стороны некоторых исследователей, которую изучающие ДП обычно приписывают зависти, [объясняется] отсутствием другой [лучшей] игрушки» [112, p.923]
С системной позиции феномен ДП может быть рассмотрен как электрофизиологическое описание процесса рассогласования. Действительно, исходя из системной концепции детерминации активности нейрона (см. выше), он не реагирует на входную импульсацию, а активируется, подготавливая будущий приток и подготавливаясь к нему. Можно полагать поэтому, что «незапланированный», «неожиданный» приток вызовет рассогласование и, следовательно, активность нейрона, направленную на устранение этого рассогласования. Если так, то, по-видимому, искусственная тетаническая электростимуляция служит мощным фактором рассогласования. И повышенная активность клетки, сохраняющаяся при тестировании – отражение данного рассогласования. Поэтому логично думать, что ДП, полученная в подобных условиях, имеет артефактную природу.
В пользу связывания ДП с процессом рассогласования свидетельствуют, кроме теоретических соображений, и экспериментальные данные. Обнаружено явное соответствие ДП тем процессам, которые имеют место в патологии при стойких отклонениях свойств метаболической среды. С использованием метода регистрации отдельных нейронов задних рогов спинного мозга крыс с экспериментально вызванным воспалением (моноартритом) показано, что стимуляция С-волокон у них значительно легче вызывает ДП, чем у здоровых животных. Авторы считают, что «механизмы, лежащие в основе ДП, сходны с теми, которые обусловливают сенситизацию при периферическом воспалении» [159, p.51].
Итак, хотя тетанизирующие экспериментаторы, не планируют вызвать рассогласование, именно его они вызывают. А рассогласование есть, как аргументировалось выше, первый этап научения, формирования новой памяти. Именно в этом смысле я и рассматриваю ДП как феномен, имеющий, возможно, отношение к механизмам научения и памяти. Имеющий вовсе не потому, что моделирует «повышение эффективности синаптической проводимости». А имеющий «возможно» потому, что неизвестно, обладает ли получаемое при индукции ДП рассогласование теми свойствами, которые присущи естественному рассогласованию при научении. Поэтому в названии настоящего раздела присутствует вопрос.
Shors T.J. и Matzel L.D [145] на основе подробного анализа экспериментальных данных о свойствах ДП пришли к заключению о несоответствии свойств ДП тем, которые должны были бы быть, если бы ДП обеспечивала сохранение долговременной памяти. В качестве наиболее серьезного сомнения при рассмотрении ДП в качестве механизма долговременной памяти авторы рассматривают ее недостаточную длительность. Да, ДП длится часы и дни, но достаточно ли это долго, чтобы обеспечить поддержание долговременной памяти? - спрашивают авторы. (К сомнениям авторов в достаточности длительности ДП для рассмотрения ее в качестве механизма долговременной памяти добавлю, что и та длительность, которая получается в подавляющем большинстве экспериментов с тетанической стимуляцией, с подведением БАВ и последующим тестирующим раздражением и которую обсуждают критики, возможно больше периода повышенной возбудимости, имеющего место в натуральных условиях (см. далее о работе [43]). Длительность может быть повышена потому, что у индивида не существует фило- или онтогенетически приобретенной памяти об устранении рассогласования, возникающего при электрической стимуляции мозга или инъекции значительных объемов БАВ.) Авторы приходят к выводу о том, что ДП - механизм, имеющий отношение не к поддержанию долговременной памяти, но к начальному периоду ее формирования и связан с механизмами пробуждения и внимания. Shors T.J. и Matzel L.D решают проблему не так, как предлагается здесь, но и они связывают ДП именно с инициацией процессов научения.
Для потока исследований ДП более шести лет, прошедшие с момента опубликования обзорно-теоретической статьи Shors T.J. и Matzel L.D, - большой срок. Однако и в настоящее время можно считать неустраненными противоречия между постулируемым механизмом долговременной памяти и свойствами ДП. Подтверждения того, что ДП моделирует механизм, лежащий в основе сохранения памяти за счет изменения эффективности синаптической проводимости, «отсутствуют сегодня», как и 10 лет назад, - констатируют J. W. Lichtman и J.R. Sanes [110, p.928]. Но «когда люди ищут что-нибудь, они часто это находят, даже если этого нет» [110, p. 929], - замечают они.
На фоне множества работ, проанализированнах Shors T.J. и Matzel L.D, как успешную находку можно оценить исследование W.C. Abraham et al. [43]. Авторы смогли подобрать условия, при которых ДП в гиппокампе крыс поддерживалась в течение нескольких месяцев. С традиционных позиций эти данные можно рассмотреть как подтверждение того, что ДП-подобный механизм может участвовать в поддержании долговременной памяти. (Правда, при этом возникают опредленные противоречия между тем, что феномен особенно длительной ДП обнаружен именно в гиппокампе, и представлением об участии гиппокампа лишь в начальных этапах поддержания памяти.)
Но с системных позиций можно рассмотреть данные W.C. Abraham et al. так, что они, в отличие от множества других авторов, лишь смоделировали такую ситуацию, в которой рассогласование, индуцированное тетанизацией, не устраняется сравнительно долго. Если принять последнюю точку зрения, то следует думать, что авторы получили рассогласование, длительность которого существенно превышает ту, которая имеет место в натуральных условиях. Это бы означало, иначе говоря, что с длительностью соответствующих процессов научения более сопоставимы данные авторов, демонстрирующих сравнительно короткоживущую ДП, чем результаты W.C. Abraham et al. Интересно, что W.C. Abraham et al. сами высказывают вполне обоснованное, с моей точки зрения, сомнение в том, что «механизмы памяти в реальном мире требуют такой стабильной пластичности»[43, p.9632].
В данной работе показано также, что помещение животных в обогащенную среду после индуцирования ДП, приводит к редукции последней. (Можно предположить, что процессы, индуцируемые помещением в обогащенную среду, накладываются на уже развернутое рассогласование, вызванное индукцией ДП, и облегчают поиск новых способов согласования клеток. То есть этот поиск происходит в контексте устранения «естественного» рассогласования.) Имея в виду упомянутую редукцию, W.C. Abraham et al. заключают, что «в норме при воздействии реальной и изменяющейся среды ДП в гиппокампе … может быть существенно менее длительным» (short-lasting) [43, p.9632].
Исходя из предположения о ДП как электрофизиологическом описании процесса рассогласования, можно думать, что даже в тех случаях, когда речь идет о длительности, измеряемой не месяцами, а днями, то, хотя такая длительность недостаточна для рассмотрения ДП в качестве феномена, отражающего механизмы долговременной памяти, она в части случаев достаточна для того, чтобы рассмотреть ее как отражение затянутого рассогласования, ведущего к гибели нейронов. Проще говоря, логика предлагаемых здесь представлений допускает и даже предполагает возможную связь ДП с гибелью нейронов. В литературе существуют данные, свидетельствующие в пользу этого предположения.
Рассогласование, как уже отмечалось выше, может быть вызвано не только электрической стимуляцией, но и введением БАВ. Введение агониста метаботропных рецепторов mGluR: ACPD [1S,3R]-1-aminocyclopentane-1,3-dicarboxylate in vitro и in vivo индуцирует развитие медленно начинающейся (slow-onset) ДП в CA1, выражающейся при внутриклеточной регистрации в увеличении амплитуды ВПСП [58]. В лаборатории K.G. Reymann [113] выясняли, «каковы последствия этого феномена для клеток интактного гиппокампа». Авторы пришли к выводу о существовании явной связи «этой синаптической пластичности с патологическими процессами в гиппокампе in vivo» [113, pp.487, 493]. ДП, которая длилась более 4 часов и индуцировалась превышающей пороговую концентрацией ACPD (20 nmol), влекла за собой выраженную гибель клеток CA1. Концентрации, которые не индуцировали ДП, не оказывали (или оказывали незначительный) нейротоксический эффект. Авторы отмечают, что гибель клеток может иметь место и в экспериментах других авторов, в которых индуцируется ДП.
Выдвижение и селекция гипотез при научении
Выше говорилось о том, что рассогласование устраняется путем проб и поиска новых путей достижения результата поведения. Может ли произойти селекция нейронов, их специализация и фиксация новой системы в памяти без реализации внешне наблюдаемого («внешнего») поведения? Если да, то как быть с положением о том, что новая интеграция фиксируется достигнутым результатом?
Описание соотношения организма со средой в новой ситуации как процесса, включающего выдвижение и селекцию гипотез, было в яркой форме представлено К.R. Popper [127]. D. Dennett [73], рассматривая гипотетическое «скиннеровское» существо, осуществляющее отбор одного удачного акта из ряда путем реализации последовательных проб «вслепую», отмечает, что каждая из таких проб может привести к гибели существа. Автор справедливо считает более эффективным способом формирования нового опыта соотношения организма со средой предварительную, «внутреннюю» селекцию актов, которую осуществляет «попперовское» существо. «Внутренняя» селекция делает возможной ситуацию, в которой, по словам К.Поппера, вместо нас гибнут наши гипотезы. Плодотворность идей К. Поппера о формировании и селекции гипотез для разработки представлений о механизмах научения отмечал П.В. Симонов [30]. В настоящее время эти идеи эффективно используется при формулировке новых концептуальных схем в когнитивных науках.
Нарастание способности совершения проб и ошибок «в уме», без реализации их во «внешнем» поведении, рассматривалось Л.В. Крушинским [23] в качестве показателя развития поведения в филогенезе. В области практической деятельности человека (спортивной, музыкальной и т.д.) реальность этой стадии формирования памяти уже давно была осознана. Анализ процесса освоения нового музыкального произведения или обучения новому акробатическому движению приводит, соответственно, к заключениям о том, что каждый пассаж должен быть «готов психически», прежде чем он будет испробован на рояле [18], и что чем лучше занимающийся представит изучаемое акробатическое движение [22], тем быстрее будет происходить процесс обучения.
Следует полагать, что выдвижение акта-гипотезы и его тестирование во «внутреннем плане», т.е. проверка гипотезы на соответствие сложившейся к данному моменту структуре памяти индивида (пробная организация совместной активности новой совокупности нейронов), занимают определенный временной интервал. Величина этого интервала зависит от многих обстоятельств: степень новизны гипотезы, сложность уже имеющегося у индивида опыта и др. Основываясь на многочисленных данных, показывающих значительное сходство паттернов мозговой активности при воображении или наблюдении действия и при реальном его осуществлении [83,98], можно допустить, что составы активированных нейронов при тестировании гипотезы во «внутреннем» и затем во «внешнем» планах существенно, хотя, конечно, не целиком, перекрываются.
В результате тестирования фиксируется новая интеграция и инициируется изменение структуры памяти. В литературе имеются данные, которые свидетельствуют в пользу возможности подобной фиксации [128]. В экспериментах с регистрацией активности нейронов у обезьян показано, что характеристики импульсации существенно изменяются уже на этапе, когда животное нашло правильное решение (в следующих после этого изменения реализациях поведения животное с вероятностью, равной единице, начинает действовать безошибочно), но еще ни разу не проверило его реализацией «внешнего» поведения, завершающегося подачей пищи. Сходные закономерности, естественно, могут быть выявлены и у людей, формирующих новых поведенческий акт. Так в экспериментах с испытуемыми, участвующими в стратегической игре (крестики-нолики), до первой реализации нового для игрока хода обнаруживаются реорганизации поведения: время выбора ходов и, как предполагается, число актуализируемых в игре связанных элементов памяти изменяется [2].
Таким образом, можно считать, что фиксация новой интеграции не обязательно требует достижения результата внешне наблюдаемого поведения. Но это, однако, не означает, что нарушается одно из основных положений теории функциональных систем: о результате как системообразующем факторе. В качестве системообразующеого фактора в этом случае выступает результат тестирования гипотезы во «внутреннем» плане.
Нет оснований думать, что консолидация памяти, лежащая в основе описанной фиксации интеграции, происходит мгновенно. В связи с этим возникает вопрос, отличаются ли стадии консолидации памяти в этом случае и, если отличаются, то как, от стадий консолидации памяти, формирующейся после реализации результативного «внешнего» поведения. Получение ответа на этот вопрос важно для понимания закономерностей формирования памяти.
В части случаев вновь сформированная интеграция как таковая не предполагает реализацию специального поведения для своего тестирования во «внешнем» плане; многие «внутренние действия» не «подлежат последующей экстериоризации» [20, с.171]. Подобные зафиксированные интеграции могут входить, по-видимому, в состав «семантической памяти».
В других случаях прошедший «внутреннее» тестирование акт-гипотеза формируется для того, чтобы совершить то или иное конкретное «внешнее» поведение (внешнее тестирование). Здесь после внутреннего тестирования могут иметь место минимум два исхода: такое поведение будет реализовано или нет. В первом случае, как и в
Date: 2015-10-18; view: 663; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |