Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Биосинтез нуклеиновых кислот и белков (матричные биосинтезы)
Первичную структуру важнейших биополимеров - белков и нуклеиновых кислот - можно сравнить с буквенной записью: и в том, и в другом случае имеется не произвольное, а строго определенное, «имеющее смысл» чередование элементов - мономеров или букв. На этом основании нуклеиновые кислоты и белки называют информационными молекулами. Чтобы получить такие молекулы, недостаточно смешать мономеры и обеспечить условия образования пептидной или фосфоди- эфирной связи, необходима еще программа, определяющая последовательность присоединения разных мономеров к растущей цепи полимера. При биосинтезе новых молекул нуклеиновых кислот и белков носителями такой программы являются нуклеиновые кислоты; в этой роли их называют матрицами. Матрица в ходе матричного синтеза не расходуется и может использоваться многократно; в этом отношении она сходна с катализатором.
Различают три основных типа матричных биосинтезов:
1) биосинтез ДНК (репликация ДНК) с использованием в качестве матрицы уже существующих молекул ДНК;
| небольшое число нуклеотидных пар (до 20-30) и чередуются с неспира- лизованными участками (рис. 2.5): |
|
| •—т |

|
2) биосинтез РНК на матрице ДНК (транскрипция);
3) биосинтез белков с использованием в качестве матрицы РНК (трансляция).
Репликация ДНК
Структура двойной спирали ДНК позволяет представить простой механизм репликации: две цепи ДНК, образующие спираль, сначала раскручиваются за счет разрыва водородных связей между комплементарными основаниями, цепи расходятся, а затем каждая одноцепочечная половина молекулы ДНК достраивается до целой двухцепочечной молекулы. При этом каждая цепь служит матрицей, к которой путем спаривания оснований подстраивается комплементарная цепочка. При этом свободные нуклеотиды поступают из клеточных органелл (рибосом), фермент ДНК-полимераза связывает их в фосфатно-пептидный остов, а азотистые основания этих нуклеотидов комплементарно связываются с матричными азотистыми основаниями:
A-T-A-A-G-C-T-C
• ••• а •..
• • • •, •..
T-A-T-T-C-C-A-G A-T-A-A-G-C-T-C
• ••• а •••
• • • •. •..
• • • •. •.,
T-A-T-T-C-C-A-G
Таким образом, из каждой исходной молекулы ДНК получаются две копии с идентичной структурой, т. е. образование пар оснований составляет химическую основу считывания биологической информации, закодированной в нуклеотидной последовательности.
| ^ A-T-A-A-G-C-T-C ■ A-T-A-A-G-C-T-C • •••. •.. • •••. •.. T-A-T-T-C-C-A-G \ T-A-T-T-C-C-A-G |
ДНК является носителем генетической информации. Участок ДНК, несущий информацию об одной полипептидной цепи, называется геном. Каждая молекула ДНК содержит множество разных генов, кодирующих синтез определенного белка. При делении «копии» ДНК расходятся по двум дочерним клеткам, каждая из которых вследствие этого будет иметь одну и ту же информацию, которая находится в «материнской» клетке, а значит и одинаковые наборы генов. Но непосредственного участия в синтезе белков ДНК не принимает. Она содержится в хромосомах ядра и отделена ядерной мембраной. К рибосомам посылается несущий информацию посредник -информационная РНК. Как это происходит? Для этого служат два других типа матричных биосинтезов - транскрипция и трансляция.
Биосинтез РНК (транскрипция)
Синтез и-РНК происходит в присутствии ДНК, выполняющей роль матрицы (матрицей служит одна из цепей ДНК). Синтез РНК происходит в направлении от 5'-конца к З'-концу. Все синтезированные молекулы и-РНК имеют структуру, комплементарную матрице (т. е. одной из цепей ДНК). Транскрипция катализируется ферментом РНК- полимеразой. Фермент присоединяется к матрице не в любом ее месте, а в специальных участках, называемых промоторами: в этих местах молекулы ДНК есть последовательности нуклеотидов, узнаваемые РНК- полимеразой. Связывание РНК-полимеразы с промотором приводит к локальному расхождению нуклеотидных цепей в этом участке; одна из цепей служит матрицей. Наращивание молекулы РНК происходит в результате перемещения РНК-полимеразы вдоль ДНК путем присоединения очередного рибонуклеотида, комплементарного тому дезоксирибо- нуклеотиду ДНК, который в данный момент находится в области активного центра РНК-полимеразы. В участке ДНК, где заканчивается ген, имеется последовательность нуклеотидов (терминирующий кодон), достигнув которого РНК-полимераза и синтезированная РНК отделяются от ДНК. Таким образом получаются отдельные молекулы РНК, каждая из которой содержит информацию одного гена или группы генов (называемой опероном), несущих информацию о структуре белка, необходимого для выполнения одной функции.
Биосинтез белка (трансляция)
Биосинтез белков отличается от других типов матричных синтезов двумя принципиальными особенностями:
1) нет соответствия между числом мономеров в матрице и продукте реакции (в и-РНК 4 разных нуклеотида, а в белке 20 разных аминокислот);
2) структура рибонуклеотидов (мономеров матрицы) и аминокислот (мономеров продукта) такова, что избирательные взаимодействия между ними, подобные образованию пар A-T, C-G, невозможны, иначе говоря, между и-РНК (матрицей) и пептидной цепью белка (продуктом) нет комплементарности.
Из этого следует, что механизм использования матрицы при синтезе белков должен быть иным, чем в случае синтеза ДНК и РНК. Если репликацию и транскрипцию можно сравнить просто с переписыванием текста, то трансляция - это дешифровка, декодирование информации об аминокислотной последовательности, закодированной с помощью нук- леотидной последовательности. Способ шифровки в нуклеиновых кислотах информации о первичной структуре белков получил название биологического кода (его называют также генетическим, нуклеотидным, аминокислотным кодом).
Биологический код - система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в и-РНК.
Один из первых вопросов, который возникает при выяснении структуры биологического кода, - это вопрос о кодовом числе, т. е. о числе нуклеотидных остатков, кодирующих включение в белок одной аминокислоты. Очевидно, что кодовое число не может быть равным единице, так как в этом случае с помощью четырех нуклеотидов можно было закодировать только четыре аминокислоты. При кодовом числе 2 количество разных нуклеотидных пар будет равно числу перестановки из четырех элементов по 2, т. е. равно 4 =16, что также недостаточно для кодирования всех аминокислот. Число разных троек нуклеотидов равно 4 =64. Это в три с лишним раза превышает минимальное число, необходимое для кодирования 20-ти аминокислот. Экспериментально доказано, что в биологическом коде кодовое число равно трем: тройку нуклеотидных остатков (триплет), кодирующих включение одной аминокислоты, называют кодоном.
Из 64 триплетов 61 используется для кодирования аминокислот, а три - UAA, UAG и UGA - обозначают конец матрицы: на этих триплетах обрывается дальнейшее наращивание пептидной цепи - терминирующие триплеты. Каждый триплет кодирует только какую-нибудь одну аминокислоту. Это свойство кода называют специфичностью, или однозначностью. С другой стороны, одна аминокислота может кодироваться двумя или большим числом (до шести) разных триплетов, т. е. код вырожден.
Путь информации от ДНК к белку представляется следующим образом:
| ДНК | ; 1 1 1 G-A-A-A-C-T.-C-G-G-A-T-G- •• нетранскибируемая цепь .. 3 • «.*••< C-T-T-T-G-A-G-C-C-ir-A-C-: • • - кодоны ДНК!!! ' |
| и-РНК | 1 1 1 1 G-A-A-A-C-U-C-G-G-A-U-G— - кодоны и-РНК | 1 1, ■ | ■. |
| Белок | ■ т! д ■, ; аминокислотная Glu-:—Thr--—Arg -:-Met —•• :!: ■ последовательность |
К настоящему времени биологический код изучен у большого количества разных организмов - от вирусов и бактерий до высших животных. Во всех случаях он оказался одинаковым. Эта универсальность кода лишний раз свидетельствует о единстве происхождения всех форм жизни на Земле.
Ошибка в биологическом коде приводит к различным болезням. Например, у здоровых людей в гене, несущем информацию о b-цепи гемоглобина, триплет GAA или GAG, стоящий на шестом месте, кодирует глутаминовую кислоту. У больных гемофилией второй нуклеотид в этом триплете заменен на U, а GUA или GUG кодирует аминокислоту валин.
Мутации
Изменение структуры ДНК, передающееся последующим поколениям, называется мутацией. На молекулярном уровне мутация - это изменение нуклеотидной последовательности ДНК. В какой-то степени мутация - это спонтанный процесс, которому ДНК подвергается постоянно. Причины мутаций различны:
1) связана с возможностью существования нуклеотидных оснований в двух таутомерных формах; предполагают, что в случае один на миллион возможно образование пар оснований и с таутомером;
2) действие продуктов нормального метаболизма клетки (перок- сиды, азотистая кислота) - являются мутагенами, т. е. веществами, способными индуцировать химические мутации ДНК;
3) радиация. ДНК интенсивно поглощает УФ-излучение и при достаточной дозе это излучение убивает большинство клеток, а остальные подвергаются очень глубокой мутации (водородные связи заменяются на ковалентные). Однако каждая клетка имеет механизм для восстановления ДНК: с помощью ферментов она заменяет поврежденный фрагмент.
Мутации изменяют генетический материал клетки: возникают либо болезни, либо прекращается выработка какого-либо белка, либо просто активность белка уменьшается.
Спонтанные мутации появляются в результате ошибок репликации ДНК, неправильного формирования комплементарных пар оснований, структурных искажений ДНК и вследствие перемещения подвижных генетических элементов в процессе роста и размножения популяции бактерий. Спонтанные мутации могут обусловливать благоприятные и неблагоприятные генетические изменения. Вероятность возникновения определенных мутаций в расчете на одну клетку и на одну генерацию называют частотой мутирования. При высоких скоростях роста она постоянна, и ее обычно определяют для клеток в экспоненциальной фазе роста при оптимальных условиях среды. В фенотипе проявляются не все мутации. Непроявленные мутации называются молчащими. У мутанта может произойти обратная мутация, или реверсия, в результате которой восстановятся свойства дикого штамма. Об истинной обратной мутации говорят лишь в тех случаях, когда вторая мутация точно восстанавливает исходный генотип, если же восстанавливается только фенотип, то говорят о вторичной реверсии, или супрессорной мутации. Супрессорные мутации могут происходить как в исходном гене, так и в каких-либо других участках хромосомы (интрагенные и экстрагенные супрессорные мутации).
Индуцированные мутации возникают под влиянием внешних факторов, которые называют мутагенами. Мутагены бывают физическими (УФ-лучи, у-радиация), химическими (аналоги пуриновых и пиримиди- новых оснований, например, 2-аминопурин, азотистая кислота и ее аналоги, алкилирующие агенты и др.) и биологическими (транспозоны) (см. табл. 2.4).
По протяженности повреждений мутации бывают точечными, когда повреждения ограничиваются одной парой нуклеотидов, и протяженными (аберрации). Мутации разделяют на хромосомные, обусловливающие появление нового признака при изменении двух и более участков хромосомы, и генные, обусловленные появлением нового признака при изменении гена. В этом случае может наблюдаться модификации оснований (изменения отдельных нуклеотидов), выпадение нескольких пар нуклеотидов (делеции), перемещение группы нуклеотидов в пределах хромосомы (транспозиция), разрыв путем вставки посторонней ДНК (инсерция) или добавление нуклеотидных пар (дупликация) и деформация спирали ДНК. Для точечных мутаций частота реверсий довольно высока, в то время как для аберраций реверсии не характерны.
Первичный эффект мутагенного фактора не обязательно ведет к истинной мутации. Новый фенотип проявляется только тогда, когда измененный ген начнет функционировать. С помощью различных методов удается накапливать и выделять мутантов с разного рода дефектами: с нарушением процессов транспорта или использования субстрата, с дефектами промежуточного обмена, с повышенной чувствительностью к температуре и т. д.
Теоретически, мутации, вызванные радиацией, химическими веществами или другими факторами, могли бы привести к вымиранию бактериальной популяции, однако в любой живой клетке существуют биохимические механизмы, способные полностью или частично восстанавливать исходную структуру ДНК. Совокупность ферментов, катализирующих реакции коррекции повреждений ДНК, составляют системы репарации, которые принципиально различаются по биохимическим механизмам восстановления генома. Известны три основных механизма коррекции дефектов ДНК:
1. Непосредственная реверсия от поврежденной ДНК к исходной структуре.
2. Эксцизия («выпадение») повреждений с последующим восстановлением исходной структуры.
3. Активация механизмов, обеспечивающих устойчивость к повреждениям.
Реверсия повреждений ДНК. К механизмам прямой реверсии повреждений ДНК относится световая репарация, или фотореактивация (исправление деформации ДНК под действием УФ-лучей). Световая репарация осуществляется несколькими ферментами: фотолиазой (расщепляет тиминовый димер и восстанавливает целостность соседних ти- миновых оснований), О6-метилтрансферазой (удаляет О6-метильную группу из остатков гуанина после действия метилирующих агентов), ДНК-пурин инсертазой (осуществляет встраивание утерянного при мутации основания в апуриновый сайт), ДНК-гликозилазой (удаление дефектных оснований). Все эти процессы происходят в один этап под действием конкретного фермента и безошибочно восстанавливают исходную структуру ДНК.
Системы эксцизионной репарации удаляют неправильно спаренные или поврежденные основания из ДНК и синтезируют новую последовательность ДНК, замещающую их. Место повреждения распознает эндонуклеаза, расщепляющая цепь ДНК вблизи дефекта, фрагмент удаляется, а дефект восполняется при помощи ДНК-полимеразы, которая проникает в брешь и встраивает в нее отсутствующие нуклеотиды, используя неповрежденную цепь ДНК в качестве матрицы. ДНК-лигаза ковалентно связывает 3'-конец вновь синтезированного участка ДНК с цепочкой. Поскольку эта система репарации основана на ресинтезе нук- леотидной цепи на базе неповрежденной матрицы, она также является практически безошибочной.
Репарационные механизмы устойчивости к повреждениям ДНК. Кроме механизмов исправления повреждений, клетки имеют возможность «обойти» вызванную повреждениями блокаду репликации ДНК, например, путем репарации в процессе рекомбинации.
В биохимической технологии явление мутации важно с двух точек зрения:
1. С помощью мутаций можно изменить природу микроорганизма, чтобы от стал более полезным для какой-либо определенной цели. Например, основные успехи в производстве пенициллина были достигнуты благодаря получению высокопродуктивных мутантов исходного штамма плесени Penicillium путем его облучения ультрафиолетовым светом, что привело к увеличению выхода пенициллина (нужного продукта метаболизма) на несколько порядков (с 20 мг/л до 7 г/л). В дальнейшем при использовании других мутагенов (химических веществ, вызывающих мутации) производство пенициллина удалось увеличить до 20 г/л.
Таблица 2.4
Наиболее часто применяющиеся химические агенты для получения мутагенного эффекта
| Аналоги гетероциклических оснований си н бензимидазол бензоксазол СО бензтиазол | Включаются в ДНК вместо обычных гетероциклических оснований |
| Азотистая кислота HNO2 | Проводит дезаминирование пуриновых и пири- мидиновых оснований ДНК |
| Алкилирующие агенты (хлоруксусная кислота, хлористый ацетил, диметилсульфат, иприт и др.) | Модификация пуриновых оснований, грубое нарушение их структуры, например, образуют дисульфидные мостики между цепями ДНК, препятствующие ее разделению при репликации |
2. С другой стороны, мутации могут создавать в биотехнологии и ряд затруднений. Для успеха микробиологического промышленного процесса часто необходимы чистые штаммы микроорганизмов, обладающие хорошо известными характеристиками. Вместе с тем нельзя исключать возможность мутаций в таких культурах, поэтому необходима регулярная проверка их генетической гомогенности.
Адапторная функция т-РНК
Между аминокислотами и нуклеотидами (или триплетами нуклео- тидов) невозможны комплементарные взаимодействия по типу образования нуклеотидных пар А...Т, C...G, A...U. Поэтому было сделано
предположение о существовании молекул-адапторов, каждая из которых может взаимодействовать с определенным кодоном, с одной стороны, и с определенной аминокислотой, с другой стороны. В 1957 г. эти молекулы были обнаружены. Ими оказались транспортные РНК (т-РНК). Очевидно, что для адаптирования 20-ти разных аминокислот к соответствующим им кодонам нужно 20 разных т-РНК. Эти т-РНК обозначают следующим образом: т-РНКА1а т-РНКН18 и т. д. (аланиновая т-РНК, гистидиновая т-РНК и т. д.). Однако, поскольку код вырожден, число разных т-РНК больше 20 (не меньше числа кодонов, имеющих смысл, т. е. не меньше 61-го).
Взаимодействие т-РНК с аминокислотами - ферментативный процесс, приводящий к образованию ковалентной связи между аминокислотой и т-РНК, катализируются эти реакции аминоацил-т-РНК- синтетазами. Такие соединения называют аминоацил -т-РНК (аа-т-РНК). Аминокислота присоединяется к З'-концу нуклеотидной цепи т-РНК (где имеется последовательность А - С - С, общая для всех т-РНК), при этом образуется сложноэфирная связь за счет карбоксильной группы аминокислоты и гидроксильной группы концевого остатка адениловой кислоты в т-РНК:
цитозин
--- CH2
Hwi
OH
| З'-конец т-РНК
о^=р—о
I I цитозин
O- CH2
нОн
OH
о^=р о
I I аденин
о- ch2
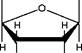 он о
он о
|
I н
о^=С----------- C------ R
| аминоацильный nh2 остаток
Существует не менее 20-ти разных аминоацил-т-РНК. Каждый из этих ферментов катализирует реакцию только одной из 20-ти аминокислот с т-РНК, соответствующей этой аминокислоте. Например, аланил-т- РНК-синтетаза катализирует реакцию аланина с аланиновой т-РНК:
Аланин + т-РНКА1а + АТФ = Ала-т-РНКА1а + АМФ + Н4Р2О7 Таким образом, аминоацил-т-РНК-синтетазы должны иметь в активном центре участок, комплементарный одной из аминокислот, и участок, комплементарный какой-то части молекулы одной из т-РНК. Именно вследствие такой субстратной специфичности каждая из ами- ноацил-т-РНК-синтетаза «узнает» и «выбирает» из смеси 20-ти аминокислот и нескольких десятков т-РНК определенную пару (аминокислоту и соответствующую ей т-РНК), и соединяет эту пару. Взаимодействие аа-т-РНК с кодоном и-РНК обеспечивается тем, что в одной из петель молекулы т-РНК имеется триплет нуклеотидов, комплементарный какому-нибудь кодону. Такой триплет называют антикодоном. Образование аа-т-РНК можно сравнить с изготовлением двойного шрифта, например, для перевода знаков азбуки Морзе в знаки буквенного алфавита:
| Phe |
| Phe-т-РНК |
к_____________ о___________ д___________
А - А - А антикодон U - U - U кодон (и-РНК)
Роль матричной РНК
Располагая двойным шрифтом, легко прочитать текст, записанный азбукой Морзе. Достаточно расставить шрифт на телеграфной ленте соответственно знакам азбуки Морзе. Роль и-РНК в трансляции аналогична роли телеграфной ленты в этом примере: аа-т-РНК присоединяется антикодонами к соответствующим кодонам и-РНК, в результате чего аминокислотные остатки оказываются расположенными в той последовательности, в какой расположены кодоны в и-РНК. Теперь остается лишь соединить аминокислотные остатки пептидной связью, чтобы получилась пептидная цепь (белок) с определенной первичной структурой. Таким образом, последовательность кодонов и-РНК коллинеарна последовательности аминокислотных остатков в соответствующем белке. Эта схема отражает лишь принципиальный механизм перевода нук- леотидной последовательности (точнее, последовательности кодонов) в аминокислотную последовательность.
Date: 2015-09-24; view: 1232; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |