Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Загальне рівняння фотосинтезу та походження кисню
Датою відкриття процесу фотосинтезу вважають 1771 рік. Англійський вчений Д. Прістлі звернув увагу на зміну складу повітря внаслідок життєдіяльності тварин. У присутності зелених рослин повітря знову ставало придатним як для дихання, так і для горіння. В подальших роботах учених (Й. Інгенхауз, Ж. Сенеб'є, Н. Соссюр, Ж. Бусенго) було встановлено, що зелені рослини із повітря поглинають вуглекислий газ, із якого за участі води на світлі утворюється органічна речовина. Саме цей процес у 1877 р. німецький вчений В. Пфеффер назвав фотосинтезом. Величезне значення для розкриття суті фотосинтезу мав закон збереження енергії, сформульований Р. Майєром (1845). Згідно з цим законом, енергія, яку використовують рослини, — це енергія Сонця, яку в процесі фотосинтезу рослини перетворюють на хімічну енергію.
Загальне рівняння фотосинтезу не досить точно передає суть цього процесу, адже, згідно з цим рівнянням, можна припустити, що частина виділеного О2 походить від СО2, тоді як насправді весь кисень утворюється з води в ході фотолізу:
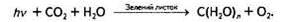
Тому доцільніше розділити його на складові частини:
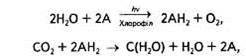
де А — акцептор електронів і протонів.
Згідно з цим рівнянням, в основі суті фотосинтезу лежить окисно-відновна реакція. За участі хлорофілу та енергії сонячних квантів вода фотоокиснюється, в результаті чого виділяються кисень та водень, останній і відновлює СО2 до рівня вуглеводів. Ці реакції відбуваються відповідно в світлову та темпову стадії фотосинтезу.
Впродовж сторіччя після відкриття Д. Прістлі (1771) фотосинтезу вчені з'ясовували його суть, встановили учасників і кінцеві продукти, що одержуються в результаті цього процесу.
Довгий час вважали загальноприйнятим, що вуглеводи утворюються з вуглецю та водню, а кисень виділяється з вуглекислоти. Ця гіпотеза, як виявилося пізніше, була абсолютно неправильною. Вперше в 1893 р. О. М. Бах передбачив, що асиміляція СО2 в процесі фотосинтезу нічого спільного з відщепленням кисню від СО2 не має, а пов'язана з окисно-відновним процесом, який відбувається з участю водню та гідроксилу води, причому кисень виділяється з води.
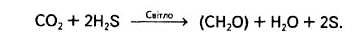
Теорію походження кисню з вуглекислого газу спростував Корнеліус Ван Ніль, який в Станфордському університеті (США) вивчав метаболізм різних фотосинтезуючих бактерій. Він з'ясував, що для фотосинтезу пурпурних сірчаних бактерій необхідний сірководень, а в результаті цього процесу всередині бактеріальних клітин нагромаджується сірка:
Після серії дослідів у 1937—1941 pp. учений зробив сміливе узагальнення: первинна фотохімічна реакція фотосинтезу полягає саме в фотодисоціації води, а не в розкладанні СО2, й запропонував сумарне рівняння фотосинтезу:
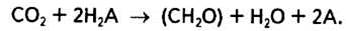
В цьому рівнянні Н2А може бути водою або іншим відновником. Отже, бактерії, здатні до фотосинтетичної асиміляції СО2, за винятком ціанел, використовують H2S, H2, СН3, а тому в процесі фотосинтезу не виділяють кисень. Такий тип фототрофного живлення дістав назву фоторедукції. Для вищих рослин і водоростей Н2А — це вода. Інакше кажучи, Ван Ніль припустив, що саме вона, а не вуглекислий газ розкладається в процесі фотосинтезу. Отже, рівняння фотосинтезу тепер матиме такий вигляд:
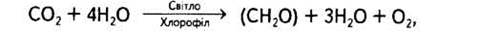
або поетапно:
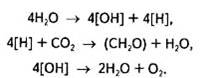
Уявлення Ван Ніля про походження кисню знайшло підтвердження в експериментах із міченим 18О2, які незалежно в 1941 р. провели С. Рубен (США) та російські вчені О. П. Виноградов і Р. В. Тейс. За допомогою мас-спектрометричного аналізу було показано, що відношення 16О2 до 18О2 в кисні, який утворюється в процесі фотосинтезу, відповідає співвідношенню їх саме у воді, а не в СО2:
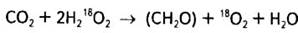
Коли ж ввести ізотоп О2 у вуглекислий газ, то рівняння набуває іншого вигляду:
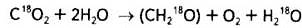
Це свідчить, що новоутворена молекула води відрізняється від тих двох молекул, що вступили в реакцію (рис. 1).
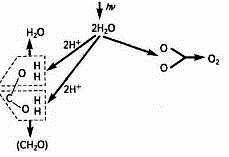
Рис. 1. Фотоліз води і виділення О2
Отже, порівняльний аналіз ізотопного складу води, вуглекислого газу та О2, що виділяється під час фотосинтетичного процесу рослиною, підтвердив спорідненість кисню фотосинтезу з водою.
Також Р. Хілл у 1939 р. продемонстрував, що ізольовані хлоропласти здатні під час освітлення розкладати воду з виділенням О2 за наявності відповідних акцепторів електронів, але без участі вуглекислого газу:
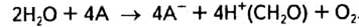
Відсутність реакції, в якій були б використані електрони, компенсується, як видно з реакції, введенням у систему акцептора електронів (А), або «реагента Хілла». Це можуть бути солі заліза (калій феріоксалат, калій фериціанід), бензохінон або природний акцептор електронів НАДФ+.
Отже, для водоростей, вищих рослин, у яких вода слугує донором електронів, підсумкове рівняння фотосинтезу матиме такий вигляд:
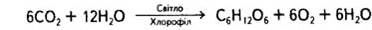
При цьому також запасається вільна енергія, величина якої для цього процесу приблизно 470 кДж/моль.
Під час фотосинтезу з простих неорганічних сполук (води та вуглекислого газу) синтезуються різноманітні органічні сполуки. В результаті відбувається перебудова хімічних зв'язків: замість С—О та Н—О виникають С—С та С—Н зв'язки, в яких електрони займають більш високий енергетичний рівень.
Дослідження свідчать також, що майже весь кисень атмосфери має фотосинтетичне походження. Отже, процеси дихання і горіння стали можливі лише після того, як виник фотосинтез. Все це й свідчить про космічне значення зеленої рослини. Підсумкова реакція фотосинтезу може мати вигляд окисно-відновних реакцій, одна з яких фотодисоціює воду, а інша — відновлює СО2 до вуглеводів, але вже без участі квантів сонячних променів: 
І дійсно, нині відомо, що фотосинтез відбувається в дві стадії — світлову та темнову.
Розділ 2. Листок як орган фотосинтезу
Особливості морфології, анатомії та фізіології листка повною мірою забезпечують здійснення його основної функції — фотосинтезу (рис. 2).
Загальна листкова поверхня рослин помірної зони, розміщених на 1 га орної площі, майже в п'ять разів перевищує земельну ділянку, а в південних районах з нормальним зволоженням — у 10... 15 разів. Різноманітна їхня форма, розташування на стеблі, фото-тропічна реакція забезпечують виконання найважливішої функції — поглинання світлових променів. Як і всі фізичні тіла, листок відбиває, поглинає та пропускає промені, що падають на нього (рис. 3).
Поверхня листка вкрита епідермісом. Клітини нижнього та верхнього епідермісу, за винятком тих, що створюють продиховий апарат, позбавлені хлоропластів. Вони лише пропускають світлові промені, отже, безпосередньої участі в фотосинтезі не беруть.
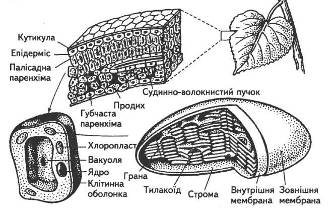
Рис. 2. Листок як орган фотосинтезу
Характерною особливістю покривної тканини листка є наявність продихів, площа яких не перевищує 1 % його загальної поверхні. Для того щоб процес фотосинтезу протікав нормально, в клітини до зелених пластид має надходити вуглекислий газ. Кількість його в атмосферному повітрі становить близько 0,03 %. Впродовж дня рослина засвоює таку кількість СО2, яка міститься приблизно в 30...60-метровому шарі повітря. Для утворення 1 г вуглеводів необхідно близько 1,47 г СО2 (така кількість його міститься майже у 2500 л повітря). Це можливо завдяки неперервному турбулентному рухові повітряних мас навкруг листків, який спричинюється нерівномірним нагріванням їх сонячними променями та вітром.
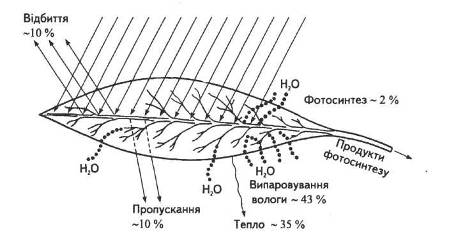
Рис. 3. Енергетичний баланс листка
Кількість води, що витрачається на фотосинтез, становить зовсім незначну частину тієї кількості, яку поглинає та випаровує рослинний організм. Дефіцит води знижує швидкість фотосинтезу, адже закриття продихів припиняє доступ вуглекислого газу до мезофілу листка. Крім того, кисень, що виділяється в процесі фотосинтетичних реакцій фотолізу води, теж потрапляє в навколишнє середовище крізь продихи. Тому закриття продихів припиняє газообмін з атмосферою, хоча при цьому ні фотосинтез, ні дихання не припиняються. Як невід'ємні структури листка, продихи взаємодоповнюють один одного, постачаючи відповідно кисень та вуглекислий газ для відповідних реакцій.
Отже, функціонування продихів регулює газообмін — надходження вуглекислого газу, виділення кисню та випаровування води. Як правило, продихів більше з нижнього боку листка, куди не потрапляють прямі сонячні промені. На верхній частині їх значно менше, що зменшує випаровування води під дією тих самих променів.
Основну ж тканину листка займає мезофіл, який буває двох типів: губчастий та палісадний (рис. 4).
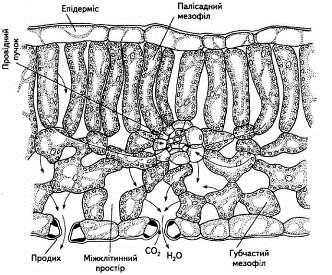
Рис. 4. Анатомічна структура листка
Найбільше хлоропластів мають клітини палісадної або, як її ще називають, стовбчастої паренхіми (кілька десятків на одну клітину). Ці клітини щільно прилягають одна до одної, на відміну від пухко розміщених клітин губчастого мезофілу. Від 15 до 20 % загального об'єму листка становить міжклітинний простір, наявність якого характерна насамперед для губчастої паренхіми. Завдяки міжклітинникам значно зростає внутрішня поверхня листка (вона в 7... 10 разів більша за зовнішню поверхню), створюється відповідне співвідношення газового складу, яке відрізняється від атмосферного, хоча міжклітинний простір і поєднується з повітрям атмосфери через продихи. Саме за рахунок міжклітинного простору СО2 постійно надходить з атмосфери, бо губчаста тканина мезофілу розміщується поблизу нижнього епідермісу, де розташовані численні продихи.
Мезофіл листка пронизаний густою сіткою судинно-волокнистих пучків, до складу яких входить ксилема, яка постачає воду та мінеральні речовини, і флоема, яка забезпечує відтік продуктів фотосинтезу.
Характерно, що прямого контакту між клітинами флоеми та кожною клітиною паренхіми немає. Це свідчить про те, що внутрішнє транспортування всього необхідного для фотосинтезу, як і відтік його продуктів на короткі відстані, здійснюється за участі симпласту й апопласту.
Для здійснення фотосинтезу в оптимальному режимі, листок має одержувати достатню кількість світлової енергії, води та вуглекислого газу.
Фотосинтез здійснюється під час безперервного надходження значної кількості світлової енергії в певні ділянки клітин. Тому в процесі еволюції рослин в клітині сформувалися спеціалізовані структури — хлоропласти (рис. 5).
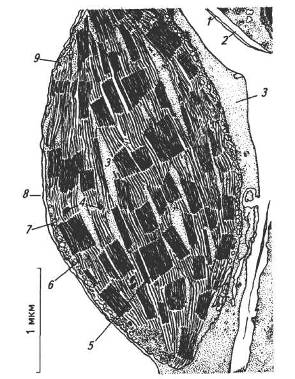
Рис. 5. Будова хлоропласта:
1 — зовнішня оболонка; 2— внутрішня оболонка; 3 — строма; 4 — краплини ліпідів; 5— грана; 6 — тилакоїди строми; 7 — крохмальні зерна; 8 — оболонка хлоропласта; 9 — рибосоми
Довжина хлоропластів коливається від 4 до 10 мкм. Кількість їх у клітині може коливатися від одного до 100 і більше. Загальна сумарна поверхня хлоропластів перевищує площу листків у десятки, навіть у сотні разів.
Хлоропласти — це напівавтономна, саморегулювальна система рослинної клітини, яка для розвитку та функціонування використовує генетичну інформацію як свого власного, так і ядерного, а можливо і мітохондріального, геномів.
У 1960 р. японський біохімік Т. Івамура виявив в одноклітинній водорості хлорелі два типи ДНК — ядерну та хлоропластну. Стало очевидним, що генетичний матеріал евкаріот локалізований не лише в ядрі, яке слугує основною базою збереження спадкової інформації, а й розсіяний по клітині в її субклітинних структурах.
Хлоропластна ДНК у вигляді подвійної спіралі має лінійну, кільце- та петлеподібну форми. Загальна кількість ДНК у хлоропласті залежно від виду коливається в межах 20... 10 • 10-15 г, а молекулярна маса — в межах 84... 104 мегадальтон. Контурна довжина кільцеподібних молекул хлоропластної ДНК у вищих рослин досягає 43...55 мм. В такій ДНК може бути близько 2 • 107 пар нуклеотидів, що відповідає наявності кількох тисяч генів.
Однак генетична інформація пластид досить обмежена. Тому багато компонентів пластид синтезуються під контролем як хлоропластної, так і ядерної ДНК. У хлоропластах є своя власна рибосомальна рРНК, а можливо, і повний набір своїх транспортних тРНК, які беруть участь в експресії геному організму. В стромі хлоропласта локалізовані дрібні рибосоми з коефіцієнтом седиментації 70S. Маючи власний спадковий апарат, власну систему, що синтезує білок, хлоропласт може контролювати синтез певних поліпептидів.
За хімічним складом основна маса хлоропластів складається (% на суху речовину) з: білків (30...60), ліпідів (20...40), хлорофілів (5...9), каротиноїдів (4...5), ДНК (0,01...0,02), РИК (О,5...3,5), мінеральних речовин (6...10). У хлоропластах зосереджено до 80 % усього заліза рослинної клітини, 65...70 % всього цинку та близько 50 % міді. Хлоропласти містять різноманітні ферменти, які забезпечують функціонування ензиматичних реакцій фотосинтезу. В них містяться всі ферменти, які беруть участь в процесі фотосинтезу (рибулозобісфосфат-карбоксилаза/оксигеназа, окисно-відновні, синтетази, гідролази та інші).
Одним із найважливіших компонентів фотосинтетичного апарату є пігментний комплекс. Прокаріоти не мають спеціалізованих фотосинтетичних органел. їхній пігментний апарат представлений в окремих мембранах, які дифузно розсіяні в клітині. Всі інші представники евкаріотів мають спеціалізовані органели — хлоропласти, де й зосереджено весь пігментний комплекс. Виявлено, що фізико-хімічні властивості пігментів найкраще реалізуються саме в хлоропластах.
Date: 2015-09-05; view: 474; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |