Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Потенциал действия
Функции нервных клеток в организме состоит в получении информации передаче ее в другие участки нервной системы, сравнение ее с информацией от других источников и, наконец, регуляции деятельности других клеток. Таким образом, живой организм представляет собой полностью электрифицированную систему. Без электричества нет жизни.
Сигналы от нервов вызывают сокращение мышечных клеток. Когда эти два типа клеток активны, возникает быстрый сдвиг мембранного потенциала в положительном направлении – потенциал действия.
Потенциалы действия можно зарегистрировать в нервных и мышечных клетках с помощью внутриклеточных электродов (слайд 4). Типичные потенциалы действия приведены на слайде 5.
Во всех этих случаях потенциал резко нарастает от отрицательных значений потенциала покоя до положительного пика, около 30 мВ, длительность потенциала действия: около 1 мс - в нервах, 10 мс - в скелетной мышце и более 200 мс - в миокарде.
Таким образом, потенциалом действия называется электрический импульс, обусловленный изменением ионной проводимости мембраны и связанные с распространением по нервам и мышцам волны возбуждения.
Потенциал действия имеет несколько фаз (слайд 6). Потенциал действия начинается очень быстрым сдвигом в положительном направлении – фаза нарастания, которая продолжается – 0,2-0,5 мс.
Во время этой фазы клеточная мембрана теряет свой нормальный заряд, или "поляризацию" поэтому фазу нарастания называют также фазой деполяризации. Как правило, деполяризация переходит за нулевую линию и мембранный потенциал становится положительным. Эта положительная фаза потенциала называется овершут. Фаза, следующая за пиком, в течение которой восстанавливается исходный потенциал мембраны в покой, называется реполяризацией.
Последний участок фазы реполяризации для некоторых видов потенциала бывает замедлен. Примерно через 1 мс после начала потенциала действия наблюдается отчетливый перегиб кривой реполяризации, следующее за ним медленной изменение называется деполяризационным следовым потенциалом. В других тканях, например нейронах спинного мозга, кривая деполяризации быстро пересекает уровень потенциала покоя, так что на некоторое время потенциал становится более электроотрицательным, чем потенциал покоя. Это явление называется гиперполяризационным следовым потенциалом.
Рассмотрим основные этапы генерации ПД, его молекулярные основы.
Почему потенциал покоя, обычно поддерживаемый на постоянном уровне нарушается до такой степени, что возникает потенциал действия? Потенциалы действия генерируются при деполяризации мембраны от потенциала до примерно – 50мВ. Механизмы развития этой начальной мы рассмотрим несколько позднее. Уровень потенциала, при котором деполяризация приводит к потенциалу действия называется порогом. При таком значении порогового потенциала заряд на мембране становится нестабильной, он нарушается посредством внутреннего механизма, который и ведет к реверсии полярности - быстрому нарастанию пика.
Опыты по исследованию потенциала действия проведены (в основном Ходжкиным и его сотрудниками) на гигантских аксона кальмара методом микроэлектродов с использованием высокоомных измерителей напряжения, а также методом меченых атомов.
В опытах по исследованию потенциала действия используются два микроэлектрода, введенных в аксон. Первый микроэлектрод - стимулирующий, на него подается импульс от генератора прямоугольных импульсов, меняющий мембранный потенциал, а мембранный потенциал измеряется при помощи второго микроэлектрода высокоомным регистратором напряжения.
Возбуждающий импульс вызывает лишь на короткое время смещение мембранного потенциала, который быстро пропадает и восстанавливается потенциал покоя. Если амплитуда импульса положительна, то такой импульс является деполяризующим и при значении большем порогового в мембране развивается процесс, в результате которого происходит резкое повышение мембранного потенциала и мембранный потенциал даже меняет свой знак - становится положительным.
После снятия возбуждения еще в течение 1-3 мс в мембране наблюдаются некоторые остаточные явления (гиперполяризационные потенциалы), во время которых мембрана рефрактерна (невозбудима).
Возникновение следующего потенциала действия в ответ на деполяризацию мембраны стимулом выше порогового может вызвать образование нового потенциала действия только после полного возвращения мембраны в состояние покоя. Причем очень важно отметить, что амплитуда потенциала действия не зависит от амплитуды деполяризующего потенциала (если он выше порогового свойства потенциала действия.
Изменение знака мембранного потенциала во время развития импульса вызвана тем, что на гребне спайка мембраны избирательно проницаема для Na+, так что отношение ионных проницаемостей становится равным РК:РNa=1:20.
Используя формулу:
 (2).
(2).
и зная РNa>>РК потенциал на мембране приближается к равновесному натриевому потенциалу 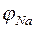 . Разность потенциалов на мембране, проницаемой только для ионов Na+, выражается формулой Нернста:
. Разность потенциалов на мембране, проницаемой только для ионов Na+, выражается формулой Нернста:
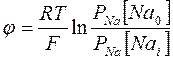 (3).
(3).
Предельное значение потенциала, вычисленное по формуле 9 равно +55мВ, что лишь немного превышает экспериментально наблюдаемые значения обращенного потенциала.
Эти выводы сохраняются, если воспользоваться для выражения мембранного потенциала более полной формулой:
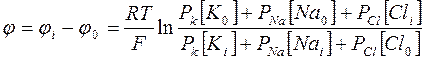 (4).
(4).
В состоянии покоя: 
РК:РNa:РСl=1:0,04:0,45, а в состоянии возбуждения на пике потенциала действия, когда вновь соблюдается условие нулевого тока через мембрану РК:РNa:РСl= 1:20:0,45.
Возможно описания изменения мембранного потенциала основанного на представлении ионных проводимостей мембраны в виде эквивалентной электрической цепи. (слайд 10). Мембранный потенциал, измеряемый по этой схеме между точками 2 и 0 (внутренняя и наружная среда клетки), создается электродвижущими силами, равными равновесными потенциалам различных видов ионов, последовательно соединенными с соответствующими сопротивлениями.
Используя условие, что в разомкнутой цепи при постоянном направлении на мембране сумма токов равна нулю, можно получить следующее выражение для мембранного потенциала:
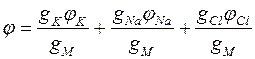 (5).
(5).
где gM – суммарная проводимость мембраны:
 (6).
(6).
Из выражения (6) видно, что значение мембранного потенциала определяется теми ионами, для которых проводимость мембраны максимальна. Зачастую уравнение (6) лучше описывает зависимость мембранного потенциала от концентрации ионов в среде, чем уравнение Гольдмана.
Процесс возбуждения развивается вследствие зависимости проницаемости ионов от мембранного потенциала. При достижении критической деполяризации, когда возрастает проницаемость мембраны для Na+, эти ионы устремляются внутрь, и вызывают дальнейшую деполяризацию мембраны.
Процесс продолжается до тех пор, пока потенциал не сместится до равновесного натриевого потенциала. В этих условиях потоки Na+ наружу и внутрь сравниваются. Затем происходит увеличение проницаемости для К+ и ионы К+ начинают выходить из клетки по градиенту своего электрохимического потенциала. В этом процессе мембрана реполяризуется. Выход К+ прекращается, когда потенциал на мембране приближается к равновесному калиевому потенциалу.
Одно из веских доказательств мембранной теории состоит в том, что проведение импульса связано с заметным возрастанием скорости движения Na+ и К+ по градиентам концентрации. Количество входящего Na+ и выходящего К+ можно определить с помощью изотопов или методом пламенной фотометрии после многократной стимуляции аксона. Результаты измерений показывают, что в гигантском аксоне каракатицы каждый импульс связан не только с входящим потоком Na+(10,3 пмоль/см2), но и с выходящим потоком этого же иона (6,6 пмоль/см2). Таким образом, суммарный вход Na+ составляет 3,7 пмоль/см2, что приблизительно соответствует потере K+ на один импульс.
Количество входящего Na+ вполне достаточно для переполяризации мембраны в период потенциала действия. Действительно, чтобы изменить напряжение конденсатора емкостью 1 мкФ на 120 мВ, необходим заряд, равный 0,12 • 10-6 Кл. Это эквивалентно 1,2 • 10-12 моль одновалентного катиона, что составляет лишь 1/3 от наблюдаемого входа Na+. Таким образом, общее количество входящего Na+ значительно превышает минимум, необходимый для изменения потенциала на мембране при возбуждении. Превышение минимума объясняется тем, что Na+ входит не только во время восходящей фазы спайка. Вход Na+ продолжается и во время ранней фазы спада, когда высока проницаемость и для Na+, и для К+. В этот период заряд, переносимый входящим Na+, с избытком компенсируется выходящим К+.
После прохождения по аксону одного спайка система возвращается в исходное состояние, за исключением того, что в клетке появился некоторый избыток ионов Na, вошедших в период деполяризации, и некоторый недостаток ионов К, вышедших на фазе реполяризации. Изменения внутриклеточной концентрации Na+ и К+, вызванные одиночным потенциалом действия, крайне малы, но могли бы оказаться значительными при многократном прохождении нервных импульсов. Для поддержания постоянного уровня внутриклеточных концентраций К+ и Na+ в клеточной мембране и существует система Na+, К+- насоса, обеспечивающая активное выведение Na+ в обмен на поступление в клетку K+, работающая за счет энергии АТФ
Литература
1. Антонов В.Ф., Черныш А.М., В.И. Пасечник и др. Биофизика. М., Владос, 2000
2. Антонов В.Ф., А.В. Коржуев Физика и биофизика. Курс лекций для студентов медицинских вузов. – М.:ГЭОТАР-Медиа, 2007 – 240 с.
3. Владимиров Ю.А. с соавт. Биофизика. М., Медицина, 1983.
4. Рубин А.Е. Биофизика: Биофизика клеточных процессов 2- том. – М. Книжный дом “Университет”, 2000 – 468 с
5. Медицинская биофизика, ред. Самойлов В.О., Л., 2004, 478c.
6. Беркинблит М.Б., Глаголева Е.Г. Электричество в живых организмах. М.: Наука, 1988. 98 с.
7. Коган А.Б. Электрофизиология. М.: Высш. школа, 1969. 368 с.
Date: 2015-09-18; view: 502; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |