Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Глава 1. Морфофункциональная организация соматосенсорной системы
1.1. Кожная рецепция. Тактильная чувствительность
Рецепторная поверхность кожи огромна (1,4—2,1 м2). В коже сосредоточено множество рецепторов, чувствительных к прикосновению, давлению, вибрации, теплу и холоду, а также к болевым раздражениям. Их строение весьма различно. Они локализуются на разной глубине кожи и распределены неравномерно по ее поверхности. Больше всего таких рецепторов в коже пальцев рук, ладоней, подошв, губ и половых органов. У человека в коже с волосяным покровом (90 % всей кожной поверхности) основным типом рецепторов являются свободные окончания нервных волокон, идущих вдоль мелких сосудов, а также более глубоко локализованные разветвления тонких нервных волокон, оплетающих волосяную сумку. Эти окончания обеспечивают высокую чувствительность волос к прикосновению. Рецепторами прикосновения являются также осязательные мениски (диски Меркеля), образованные в нижней части эпидермиса контактом свободных нервных окончаний с модифицированными эпителиальными структурами. Их особенно много в коже пальцев рук. В коже, лишенной волосяного покрова, находят много осязательных телец (тельца Мейснера). Они локализованы в сосочковом слое дермы пальцев рук и ног, ладонях, подошвах, губах, языке, половых органах и сосках молочных желез. Эти тельца имеют конусовидную форму, сложное внутреннее строение и покрыты капсулой. Другими инкапсулированными нервными окончаниями, но расположенными более глубоко, являются пластинчатые тельца, или тельца Фатера—Пачини (рецепторы давления и вибрации). Они есть также в сухожилиях, связках, брыжейке. В соединительнотканной основе слизистых оболочек, под эпидермисом и среди мышечных волокон языка находятся инкапсулированные нервные окончания луковиц (колбы Краузе) (Покровский В.М., 2011).
Кожные рецепторы можно классифицировать в соответствии с типом стимула, на который они отвечают. Основные их типы: механорецепторы, терморецепторы и ноцицепторы.
Абсолютный порог ощущения прикосновения механорецепторов в течение более 100 лет измерялся комбинированными волосками Фрея. Метод измерения разработан немецким физиологом Максом фон Фреем и до настоящего времени применяется в клинике. Современные механические стимуляторы, выполненные на основе электромагнита, обеспечивают градуированное воздействие с помощью специального стержня определенного диаметра. Таблица 1 позволяет сравнить данные о порогах тактильного восприятия, измеренных с помощью волосков Фрея, и электромагнитного стимулятора, дающего вибрации с частотой 200 Гц. Различные части тела неодинаково чувствительны к тактильным стимулам. Наиболее низкие пороги (высокая чувствительность) имеют кончики пальцев рук, губы и язык, наиболее высокие – кожа бедер, плеч, спины. Пороги постепенно возрастают от кончиков пальцев к дистальным частям руки.
Таблица 1
Пороги восприятия механической стимуляции кожи человека (ссылка
| Участок тела | Точечная стимуляция волосками Фрея, мг | Стимуляция площади в 1 см2 вибратором с частотой 200 Гц, мкм |
| Нос | 5,3 | 4,7 |
| Лоб | 7,8 | 4,2 |
| Кончик пальца | 0,07 | |
| Ладонь | 0,07 | |
| Плечо | 2,2 |
Продолжение таблицы 1
| Подошва | 0,45 | |
| Икра | 5,6 | |
| Вентральная поверхность бедра | 2,8 | |
| Грудь | 1,7 | |
| Живот | 5,2 | |
| Плечо | 1,4 |
Интенсивность тактильных раздражителей (давление), постепенно увеличивающихся над порогом, оценивается испытуемыми по-разному, что выражается различными показателями степенных функций.
Пространственное различение стимулов, измеренное с помощью циркуля, определяется как минимальное расстояние, при котором оба кончика циркуля еще ощущаются раздельно. Это так называемое одновременное пространственное различение (или двухточечный порог).
При помощи ножек циркуля с тупыми концами, устанавливаемых на кожу неодновременно, легко определяются пороги последовательного пространственного различения, т.е. минимального расстояния между двумя тактильными стимулами, при последовательном действии которых они воспринимаются как одиночный стимул (Альтман Я.А., 2009).
Механорецепторы бывают быстро или медленно адаптирующимися. К быстро адаптирующимся относятся рецепторы волосяных фолликулов волосистой части кожи, тельца Мейснера безволосой кожи ительца Пачини. Рецепторы волосяных фолликулов и телец Мейснера предпочтительно отвечают на стимулы, поступающие с частотой примерно 30-40 Гц, тогда как тельца Пачини – на стимулы с частотой примерно 250 Гц. Медленно адаптирующиеся кожные механорецепторы – это диски Меркеля и тельца Руффини. Диски Меркеля имеют точечные рецептивные поля, клетки которых расположены в базальном слое кожи ладоней и подошв. Тельца Руффини активируются при растяжении кожи, причем даже на некотором расстоянии от рецептора. Чаще встречаются в коже подошвенной поверхности стопы, а также в более глубоких тканях (например, в капсуле суставов). У всех этих рецепторов миелинизированные аксоны (Камкин А.Г., 2004).
Функции всех механорецепторов делятся на датчики прикосновения, датчики давления стимула, датчики скорости его воздействия и датчики ускорения. В таблице 2 суммируются структурно-функциональные свойства и соотношения кожных рецепторов.
Кожа содержит множество так называемых свободных нервных окончаний, на которых отсутствуют корпускулярные структуры. Афферентные волокна свободных нервных окончаний немиелинизированы и характеризуются скоростью проведения около 1 м/c. Свободные нервные окончания реагируют импульсом не более чем на один уровень интенсивности – пороговый. Поэтому они считаются пороговыми датчиками, отмечающими наличие стимула в определенном месте кожи. Свободные нервные окончания принимают участие в передаче информации о слабых движущихся по коже стимулах и имеют отношение к ощущениям щекотки и зуда.
Таблица 2
Классификация функциональных свойств кожных механорецепторов
| Адаптация к постоянному давлению | Медленная | Быстрая | Очень быстрая |
| Тип Рецепторов и их локализация | Участки кожи, не покрытые волосками | ||
| Диск Меркеля | Тельца Мейснера | Тельце Пачини | |
| Волосистые участки кожи | |||
| Тактильные диски, окончания Руффини | Рецептор волосяного фолликула | Тельце Пачини | |
| Функциональное назначение | Датчики интенсивности (рецепторы Давления) | Датчики скорости (рецепторы прикосновения) | Датчики ускорения (вибро- рецепторы) |
Ощущение прикосновения или давления (тактильные ощущения) можно вызвать только в определенных точках кожи, которые называются чувствительными тактильными точками. Наибольшее число тактильных точек и разнообразных рецепторов сосредоточено на коже кончиков пальцев, на губах и на роговице глаза.
На коже, покрывающей плечи, бедра и спину, тактильные точки встречаются редко. Для появления тактильного ощущения на внутренней стороне руки достаточно вдавить кожу на глубину 0,01 мм (10 мкм). Пороги тактильных ощущений на кончиках пальцев ниже, чем на остальной поверхности кисти, что связано с их богатой иннервацией и ее высокой плотностью.
Решающую роль в пространственном разрешении играет число афферентных волокон на 1 см2 поверхности кожи (плотность иннервации). Для человека максимальное пространственное разрешение было установлено для кончиков пальцев руки, где находятся в основном быстро адаптирующиеся рецепторы – тельца Мейснера.
Тельца Пачини – детекторы коротких механических воздействий. Они расположены глубоко в коже и в подкожной клетчатке преимущественно в пальцах, наружных половых органах, фасциях, связках, суставах и молочной железе. Многократное частое раздражение этих рецепторов вызывает ощущение вибрации. Капсула тельце Пачини состоит из множества одинаковых слоев, вложенных друг в друга наподобие матрешки. В середине матрешки находится нервное окончание. Тельце Пачини относят к рецепторам ускорения.
При воздействии механического раздражителя капсула тельца Пачини деформируется и действует на нервное окончание. В нервном окончании возникает генераторный потенциал. При многократном раздражении амплитуда генераторных потенциалов телец Пачини снижается.
Потенциалы действия нервных волокон, отходящих от телец Пачини, следуют частоте раздражения до 200-400 Гц, в интервале 400-500 Гц соответствие импульсации частоте стимула нарушается, а выше 500 Гц перестают реагировать. Люди при одинаковом раздражении кожи ладони отмечают два вида ощущений – вибрации и трепетания. Для возникновения ощущения вибрации необходимы более высокая частота и меньшая амплитуда (сила), чем для ощущения трепетания, которое появляется при более низких частотах, но большей амплитуде колебаний. Трепетание исчезает при поверхностной анестезии, так же как разряды рецепторов, реагирующих на низкие частоты вибрации. Местная анестезия поверхностных слоев кожи не уничтожает вибрационную чувствительность и ответы высокочастотных рецепторов, поскольку тельца Пачини располагаются в глубоких структурах кожи и подкожных тканей. Условием появления ощущения вибрации в коже служит одновременное вовлечение в реакцию нескольких телец Пачини, поскольку именно в этом случае импульсный разряд в афферентных волокнах становится эффективным стимулом (Альтман Я.А., 2009).
Температура тела человека колеблется в сравнительно узких пределах, поэтому информация о температуре окружающей среды, необходимая для деятельности механизмов терморегуляции, имеет особое важное значение. Терморецепторы располагаются в коже, роговице, слизистых оболочках, а также в ЦНС (гипоталамус). Выделяют два вида терморецепторов: холодовые и тепловые. Они медленно адаптирующиеся рецепторы, хотя могут ответить фазическим разрядом на быстрые изменения температуры кожи. Больше всего терморецепторов в коже лица и шеи.
Терморецепторы разделяют на специфические и неспецифические. Первые возбуждаются лишь под влиянием температурных воздействий, вторые отвечают и на механические раздражение. Рецептивные поля большинства терморецепторов локальны. Терморецепторы реагируют на изменение температуры повышением частоты генерируемых импульсов, устойчиво сохраняющимся в течение всего времени действия стимула. Повышение частоты импульсации пропорционально изменению температуры, причем постоянная импульсация у тепловых рецепторов наблюдается в диапазоне температуры от 20 до 50оС, у холодовых – от 10 до 41оС. Дифференциальная чувствительность терморецепторов высока: достаточно изменить температуру на 0,2оС, чтобы вызвать длительные изменения их импульсации.
В некоторых условиях холодовые рецепторы могут быть возбуждены и теплом (выше 45оС). Этим объясняется возникновение острого ощущения холода при быстром погружении в горячую ванну. Важным фактором, определяющим активность терморецепторов, связанных с ними центральных структур и ощущений человека, является абсолютное значение температуры. В то же время начальная интенсивность температурных ощущений зависит от разницы температуры кожи и действующего раздражителя, его площади и места приложения. Если руку держали в воде температуры 27оС, то в первый момент при переносе руки в воду, нагретую до 25оС, она кажется холодной, однако уже через несколько секунд становится возможной истинная оценка абсолютной температуры воды (Покровский В.М., 2011).
Болевые рецепторы реагируют на стимулы, угрожающие организму повреждением. Существует два основных типа ноцицепторов: Аδ-механоноцицепторы и полимодальные С-ноцицепторы. Механоноцицепторы иннервируются тонкими миелинизированными, а полимодальные С-ноцицепторы – немиелинизированными С-волокнами. Аδ-механоноцицепторы отвечают на сильное механическое раздражение кожи, например, укол иглой или щипок пинцетом. Обычно они не реагируют на термические и химические болевые стимулы, если только не были предварительно сенситизированы (повышение чувствительности афферентных волокон рецепторов). В отличие от них полимодальные С-ноцицепторы) реагируют на болевые стимулы разного вида: механические, температурные и химические (Камкин А.Г., 2004).
Тактильная чувствительность (от лат. tactilis – осязаемый, от tango – касаюсь) - ощущение, возникающее при действии на кожную поверхность различных механических стимулов. Она обеспечивает ощущения прикосновения, давления, вибрации и щекотки. К тактильным рецепторам кожи относятся тельца Мейснера, диски Меркеля, тельца Пачини, рецепторы волосяного фолликула, свободные нервные окончания поверхностных слоев кожи (рецепторы щекотки), полимодальные свободные нервные окончания. Также на прикосновение реагируют колбы Краузе и тельца Руффини, являющиеся терморецепторами (Дегтярев В.П., 2012).
Ощущение прикосновения и давления на кожу довольно точно локализуется, т. е. относится человеком к определенному участку кожной поверхности. Эта локализация вырабатывается и закрепляется в онтогенезе при участии зрения и проприорецепции. Абсолютная тактильная чувствительность существенно различается в разных частях кожи: от 50 мг до 10 г. Пространственное различение на кожной поверхности, т. е. способность человека раздельно воспринимать прикосновение к двум соседним точкам кожи, также сильно отличается в разных ее участках. На слизистой оболочке языка порог пространственного различия равен 0,5 мм, а на коже спины — более 60 мм. Эти отличия обусловлены главным образом различными размерами кожных рецептивных полей (от 0,5 мм2 до 3 см2) и степенью их перекрытия (Покровский В.М., 2011).
Ощущение прикосновения – результат стимуляции чувствительных нервных окончаний кожи и подлежащих тканей.
Ощущение давления возникает в результате деформации глубоких тканей.
Вибрационное ощущение возникает в результате быстрых повторных сенсорных стимулов, наносимых на те же рецепторы, что и рецепторы, воспринимающие прикосновение и давление (Орлов Р.С., 2010).
По функциональным особенностям тактильные рецепторы подразделяются на фазные и статические. Фазные тактильные рецепторы возбуждаются при динамическом раздражении. Они обладают высокой чувствительностью, коротким латентным периодом, быстро адаптируются. Статические тактильные рецепторы возбуждаются при постоянном раздражении одинаковой силы. Они менее чувствительны, чем фазные, имеют более длительный латентный период, медленно адаптируются (Дегтярев В.П., 2012).
Восприятие сенсорных сигналов требует тонкой дифференцировки (например, определения точной локализации на коже, минимальных градаций в интенсивности). Передача таких тактильных ощущений от специализированных сенсорных рецепторов происходит по миелинизированным нервным волокнам со скоростью от 30 до 70 м/с. Свободные нервные окончания передают сигналы со скоростью от 5 до 30 м/с. Грубые виды сигналов (например, сильное давление, прикосновение без четкой локализации и в особенности щекотание) передаются нервными волокнами малого диаметра с небольшой скоростью (так, от нервных окончаний, формирующих ощущение щекотки, сигналы идут по безмиелиновым нервным волокнам С-типа со скоростью до 2 м/с) (Орлов А.Д., 2006).
1.2. Проводящие пути соматосенсорной системы
Сенсорный проводящий путь – последовательность связанных друг с другом элементов: первичных, вторичных, третичных и высших сенсорных нейронов (рис. 1.). В передачу сенсорной информации одинаковой модальности часто вовлечено несколько параллельных сенсорных путей.
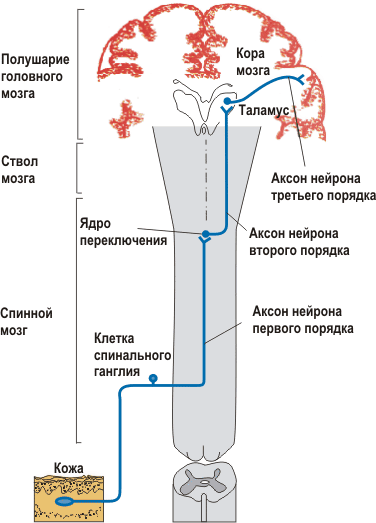
Рис. 1. Общая организация сенсорных проводящих путей. На схеме показаны нейроны первого, второго и третьего порядков.
Нейроны первого порядка в сенсорном пути – это первичные афферентные нейроны. Периферические окончания их аксонов являются сенсорными рецепторами (или получают вход от добавочной сенсорной клетки, примером которой может служить волосковая клетка улитки внутреннего уха). Таким образом, первичные нейроны реагируют на стимул, преобразуют его энергию, а затем направляют перекодированную информацию в ЦНС. Сома первичного афферентного нейрона обычно располагается в ганглии заднего спинномозгового корешка или черепного нерва.
Нейроны второго порядка находятся в спинном мозгу или в стволе мозга. Они получают сигналы от нейронов первого порядка (обычно от нескольких или даже многих) и передают информацию в таламус. В этом существенную роль играют переработка сигналов в локальных нейронных сетях и биофизические процессы в мембране этих нейронов. Аксоны нейронов второго порядка восходят к таламусу, переходя на другую сторону мозга. Сенсорная информация от одной половины тела поступает к противоположной (контралатеральной) части таламуса.
Нейроны третьего порядка находятся в каком-либо из сенсорных ядер таламуса. Информация, получаемая ими от нейронов второго порядка, перерабатывается по такому же принципу, как в последних, и направляется в кору больших полушарий.
Нейроны четвертого порядка соответствующей области сенсорной коры и высшие нейроны в той же и других ее областях производят дальнейшую переработку информации. И наконец, в одном из участков коры (пока не установленном) формируется восприятие – осознанное представление о стимуле (Камкин А.Г., 2004).
1.3. Соматосенсорная система
Кора больших полушарий человека подразделяется на 50 различных областей, называемых полями Бродманна. Сенсорные сигналы всех модальностей (кроме зрительных и слуховых) поступают в кору мозга позади центральной борозды. Передняя половина теменной доли почти полностью занята рецепцией и интерпретацией соматосенсорных сигналов. Задняя половина теменной доли осуществляет высший уровень интерпретации.
Область мозговой коры, лежащая кпереди от центральной борозды (задняя половина лобной доли) контролирует мышечные сокращения и движения тела.
В соматосенсорной коре выделены две области – соматосенсорная область I и соматосенсорная область II (рис.2.А).
Соматосенсорная кора I располагается позади центральной борозды занимая поля 1, 2 и 3 по Бродманну. В ней расположены точки восприятия различных частей тела – корковое представительство (рис.2.Б).
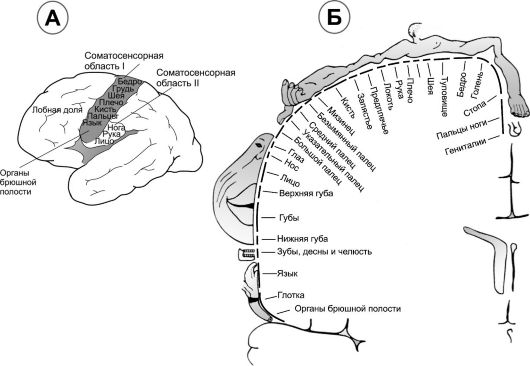
Рис. 2. Соматосенсорная кора.А. локализация соматосенсорных областей I и II на поверхности большого мозга. Б. Соматотопическое представительство различных областей тела в сенсомоторной области I (фронтальное сечение мозга на уровне постцентральной извилины)
Импульсы соматосенсорной области II поступают в эту область из ствола мозга, передающего сигналы с обеих сторон туловища и конечностей. Кроме того, сюда поступают сигналы из соматосенсорной области I, а также от других сенсорных областей (например, слуховой и зрительной коры).
Соматосенсорная кора каждого полушария получает информацию от противоположной стороны тела (за исключением небольшое количество информации, идущее от той же стороны лица). На рис.2.Б самыми большими участками коры представлены губы, затем лицо и большой палец руки. Туловище и нижняя часть тела представлены небольшими участками. Размер участка в соматосенсорной коре прямо пропорционален количеству специализированных рецепторов в каждой периферической области тела. В губах и большом пальце кисти найдено значительное количество специализированных нервных окончаний, а в коже туловища обнаружены лишь отдельные специализированные нервные окончания.
Соматосенсорная кора организована в виде функциональных единиц – колонок нейронов, перпендикулярных ее поверхности. Каждая колонка имеет диаметр от 0,3 до 0,5 мм и содержит приблизительно 10 тыс. тел нервных клеток. Каждая колонка обслуживает отдельную специфическую сенсорную модальность. Некоторые колонки отвечают за рецепторы растяжения вокруг суставов, другие – за стимуляцию волос, третьи – за дискретную локализацию точек давления на коже и так далее. Только в слое IV соматосенсорной коры, где сенсорные сигналы входят в кору, нейронов функционируют отдельно друг от друга. На всех остальных уровнях колонки взаимодействуют, осуществляя начальный анализ значения сенсорных сигналов.
После двустороннего удаления соматосенсорных областей I наблюдаются следующие изменения: теряется способность точной локализации ощущений в различных частях тела; утрачивается способность критически оценивать степень давления; исчезает способность судить о массе вещей на ощупь; теряется способность оценивать вид или форму предметов на ощупь (утрата этой способности называется астереогнозис). Болевое и температурное чувство после удаления соматосенсорной коры не претерпевает значительных изменений в отношении интенсивности или качества ощущений, но становится затруднительной их точная локализация.
Соматосенсорные ассоциативные области и их функции. Поля 5 и 7 по Бродманну, лежащие позади соматосенсорной коры, принимают участие в расшифровке информации, поступающей в соматосенсорную кору. Отсюда их название – соматосенсорные ассоциативные области. Электрическая стимуляция соматосенсорной области вызывает у человека комплекс соматических ощущений (например, чувство мяча в руке). Соматосенсорная ассоциативная область интегрирует информацию, поступающую из различных точек тела в соматосенсорную кору для расшифровки ее значения. Ассоциативная область получает сигналы из: соматосенсорной области I; вентробазальных ядер таламуса; других областей таламуса; зрительной коры; слуховой коры.
Соматосенсорной ассоциативной области вызывает у человека потерю способности узнавать на ощупь комплекс предметов и их форму на противоположной стороне. Кроме того, человек теряет чувство формы своего тела или части тела на противоположной стороне (такая комплексная потеря ощущений называется аморфосинтез) (Орлов А.Д., 2006).
Date: 2015-09-18; view: 1057; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |