Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Высвобождающая единица как самоподдерживающийся кальциевый осциллятор
В программном комплексе, разработанном на базе рассматриваемой модели, исследовалась динамика ионов Са2+ в высвобождающая единице, включающей в себя кластер 9х9 RyR-каналов. Как уже отмечалось, процессы открытия/закрытия каналов в течение кальциевого цикла носят стохастический характер, и вероятность их открытия зависит от концентраций Са2+ в люмене СР и диадном пространстве. Из этого следует, что их активность должна меняться в течение Са2+-цикла в клетке.
Данный программный комплекс позволил исследовать внутренние Са2+-«часы» и получить временные зависимости таких величин, как концентрация Са2+ в люмене (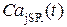 ), эффективное давления (
), эффективное давления (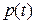 ), концентрация Са2+ в диадном пространстве (
), концентрация Са2+ в диадном пространстве (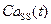 ), концентрация Са2+ в цитозоле клетки (
), концентрация Са2+ в цитозоле клетки (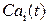 ) и число открытых каналов в кластере (
) и число открытых каналов в кластере (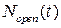 ). Подробно предположительный цикл работы кальциевых «часов» описан ранее в пункте 3.2.4 настоящей работы.
). Подробно предположительный цикл работы кальциевых «часов» описан ранее в пункте 3.2.4 настоящей работы.
На рисунке 4.9 представлены результаты численного моделирования Са2+-«часов» при следующих значениях параметров динамики 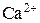 :
: 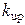 =0.0012 М·с-1,
=0.0012 М·с-1,  = 6·10-7 М,
= 6·10-7 М, 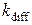 = 25·103 с-1,
= 25·103 с-1,  = 50 с-1,
= 50 с-1, 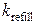 =1 с-1. Параметры концентраций буферов, учитывающихся в модели, выбирались согласно работе [80]: CQ tot = 10-2 М, CM tot = 45·10-6 М, TC tot = 31·10-6 М.
=1 с-1. Параметры концентраций буферов, учитывающихся в модели, выбирались согласно работе [80]: CQ tot = 10-2 М, CM tot = 45·10-6 М, TC tot = 31·10-6 М.
На данном этапе моделирование проводилось без учета конформационного взаимодействия между RyR-каналами в решетке (k =0). Параметры ЭК модели (2.5-2.8) выбирались такими же, как и в работе [A1]: a = 5, K = 12, 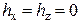 , Г = 7, n = 6,
, Г = 7, n = 6, 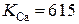 ·10-6 M. Параметры туннелирования: A = 1,
·10-6 M. Параметры туннелирования: A = 1, 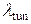 = 0.01.
= 0.01.
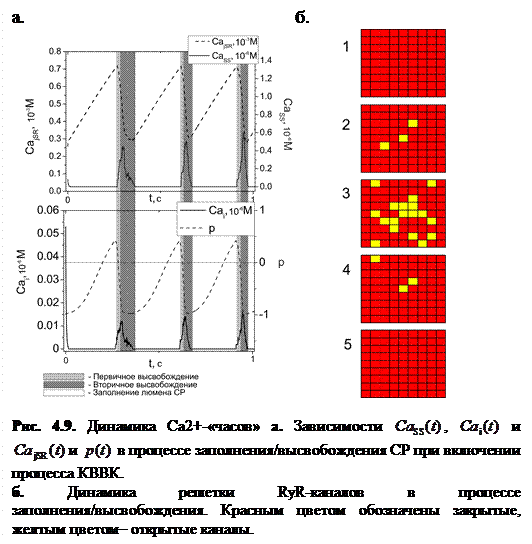
Как видно из рисунка, Са2+- высвобождающая единица действительно ведет себя как самоподдерживающийся осциллятор в определенном диапазоне динамических параметров.
Зависимость  представляет собой последовательность отдельных сигналов несинусоидальной формы с практически постоянной амплитудой (0.5±0.075 мкМ). На основании этого можно сделать вывод, что высвобождающая единица при данном значении динамических параметров ведет себя как релаксационный осциллятор. Число открытых каналов также меняется периодически. В начале эксперимента все каналы решетки закрыты (рис. 4.9б(1)), затем происходит открытие малого числа каналов, вызывая тем самым первичное высвобождение (рис. 4.9б(2)). Затем открывается большее число каналов (рис. 4.9б(3)), в связи с чем резко понижается концентрация в люмене, что стимулирует перезагрузку системы и закрытие каналов (рис. 4.9б(4, 5)). Высвобожденный кальций диффундирует сначала в цитозоль, затем возвращается в сеть ретикулума. После этого в процессе заполнения попадает обратно в люмен, и процесс высвобождения повторяется.
представляет собой последовательность отдельных сигналов несинусоидальной формы с практически постоянной амплитудой (0.5±0.075 мкМ). На основании этого можно сделать вывод, что высвобождающая единица при данном значении динамических параметров ведет себя как релаксационный осциллятор. Число открытых каналов также меняется периодически. В начале эксперимента все каналы решетки закрыты (рис. 4.9б(1)), затем происходит открытие малого числа каналов, вызывая тем самым первичное высвобождение (рис. 4.9б(2)). Затем открывается большее число каналов (рис. 4.9б(3)), в связи с чем резко понижается концентрация в люмене, что стимулирует перезагрузку системы и закрытие каналов (рис. 4.9б(4, 5)). Высвобожденный кальций диффундирует сначала в цитозоль, затем возвращается в сеть ретикулума. После этого в процессе заполнения попадает обратно в люмен, и процесс высвобождения повторяется.
Отличительной особенностью релаксационного осциллятора является независимость его кинетических параметров от начальных условий [124, 125]. Для проверки этого эффекта построены зависимости  и
и  в повторяющемся цикле работы Са2+-«часов» при различных начальных условиях (рис. 4.10).
в повторяющемся цикле работы Са2+-«часов» при различных начальных условиях (рис. 4.10).
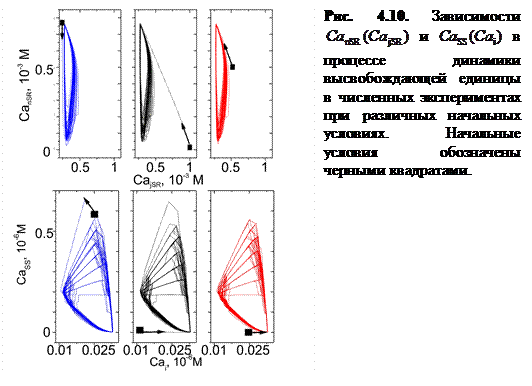
Из графиков ясно, что выбор начальных условий не влияет на характер дальнейшей динамики системы, что подтверждает устойчивость релаксационных колебаний осциллятора при данных значениях параметров.
4.2.2 Моды динамики Са2+-«часов»
Важно отметить, что в ходе моделирования при варьировании параметра 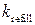 и при учете кооперативной динамики RyR-каналов были выявлены и другие типы колебаний внутренних Са2+-«часов».
и при учете кооперативной динамики RyR-каналов были выявлены и другие типы колебаний внутренних Са2+-«часов».
На рисунке 4.11 изображены зависимости концентрации Са2+ в диадном пространстве от времени при различных значениях параметра скорости заполнения люмена.
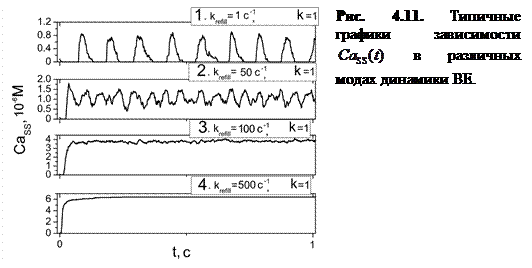
При варьировании параметра 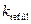 в широком диапазоне (0.05÷500 мс-1) выявлено пять основных мод динамики высвобождающей единицы, которые подробно будут описаны далее.
в широком диапазоне (0.05÷500 мс-1) выявлено пять основных мод динамики высвобождающей единицы, которые подробно будут описаны далее.
В ходе исследования зависимости частоты и амплитуды высвобождающегося Са2+ от скорости заполнения люмена и параметра взаимодействия между каналами построена – k фазовая диаграмма, совмещенная с графиками плотности, на которых выделены различные моды поведения системы в зависимости от параметров k и :
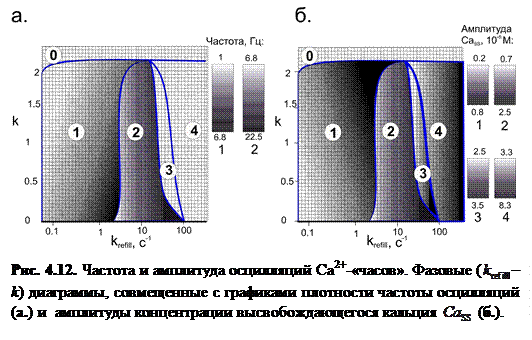
На рисунке справа от диаграмм представлены цветовые шкалы, отображающие значения частоты и амплитуды 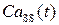 в различных модах.
в различных модах.
Мода 0: Статическое равновесие: при сильном взаимодействии между каналами происходит перенормировка ветвей конформационного потенциала в направлении стабильности закрытого состояния. Так, при k > 2.3 даже при максимальном заполнении люмена закрытое состояние остается стабильным, и в связи с этим не происходит туннелирования каналов в открытое состояние.
Мода 1: Релаксационные осцилляции. (рис. 4.11(1)). При достаточно малой скорости заполнения ( <10 с-1) система ведет себя как устойчивый релаксационный осциллятор. После завершения высвобождения уровень 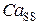 понижается до нулевого уровня вследствие низкой частоты высвобождений. В данной моде частота осцилляций варьируется в достаточно широком интервале (от 1 до 6.8 Гц), как показывает градация цвета от белого к черному на графике плотности. Также следует указать на увеличение амплитуды концентрации высвободившегося Са2+ (от 5·10-7 до 1.5·10-6 М), что видно из рис. 4.11(1).
понижается до нулевого уровня вследствие низкой частоты высвобождений. В данной моде частота осцилляций варьируется в достаточно широком интервале (от 1 до 6.8 Гц), как показывает градация цвета от белого к черному на графике плотности. Также следует указать на увеличение амплитуды концентрации высвободившегося Са2+ (от 5·10-7 до 1.5·10-6 М), что видно из рис. 4.11(1).
Мода 2: Гармонические высокочастотные осцилляции. (рис. 4.11(2)). При увеличении скорости заполнения увеличивается частота высвобождений, однако, уровень не успевает понизиться до нулевого уровня. Как видно из рисунка, осцилляции системы носят гармонический характер.
Мода 3: Динамическое квазиравновесие. (рис. 4.11(3)). При дальнейшем увеличении скорости заполнения люмена ( >50 с-1) наблюдались осцилляции высокой частоты и достаточно малой амплитуды вблизи определенного значения концентрации (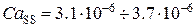 M). Данные осцилляции вследствие малой амплитуды можно рассматривать как динамическое квазиравновесное состояние системы.
M). Данные осцилляции вследствие малой амплитуды можно рассматривать как динамическое квазиравновесное состояние системы.
Мода 4: Динамическое равновесие. (рис. 4.11(4)). При большом значении параметра скорости заполнения ( >200 с-1) открывается весь кластер RyR-каналов. В этом случае наблюдается стабильное вытекание ионов 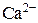 в диадное пространство с постоянной скоростью.
в диадное пространство с постоянной скоростью.
На рисунке 4.13 представлены графики зависимостей и 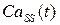 в модах 1 и 2 при различных значениях
в модах 1 и 2 при различных значениях 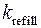 .
.
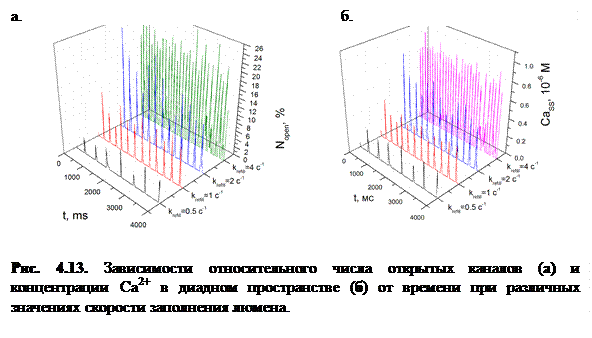
Как видно из рисунка, при увеличении происходит рост амплитуды концентрации высвободившегося Са2+. Следует отметить, что в модах 1 и 2 (оптимальный режим) наблюдается эффект увеличения амплитуды концентрации высвобождающегося с ростом частоты осцилляций (рис. 4.14). Одновременно с этим происходит увеличение амплитуды колебаний в люмене и увеличение амплитуды осцилляций числа открытых каналов 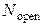 .
.
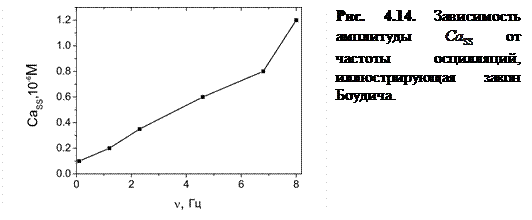
Детальный анализ [А2] показал, что данный эффект может быть объяснен как результат задержки туннелирования каналов в открытое состояние. Это явление подробно описано ранее в пункте 4.1.3 настоящей работы. При большей скорости заполнения, как было показано, эффект задержки проявляется сильнее, чем при малой скорости заполнения люмена.
Данный эффект приводит к тому, что большей скорости заполнения соответствует больший уровень концентрации 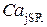 , при котором достигаются оптимальные условия туннелирования между ветвями конформационного потенциала. В связи с этим в процессе высвобождения открывается бόльшее число каналов и высвобождается бόльшее количество ионов в диадное пространство.
, при котором достигаются оптимальные условия туннелирования между ветвями конформационного потенциала. В связи с этим в процессе высвобождения открывается бόльшее число каналов и высвобождается бόльшее количество ионов в диадное пространство.
Если учесть прямую связь силы сокращений кардиомиоцитов с концентрацией высвободившегося [4, 70], то данный факт на молекулярном уровне объясняет давно известный принцип Боудича работы клеток водителей ритма «чаще-сильнее». Другими словами, внутренний СР-осциллятор в ЭК-модели может работать индивидуально как источник устойчивых сердечных сокращений.
Как уже отмечалось ранее, представления о динамике ионов Са2+ и о принципах работы внутренних Са2+-«часов» в данной работе основываются на модели Мальцева-Лакатты (ML) [87, 88]. Эта модель адекватно описывает кальциевую кинетику, но обладает рядом недостатков, в частности, пренебрежением кооперативной динамикой RyR-каналов в кластере высвобождающей единицы. Следует отметить, что ML-модель демонстрирует обратное «анти-Боудич» поведение -«часов» («выше частота, меньше амплитуда»), однако, эффект Боудича в этой модели воспроизводится только с учетом взаимодействия внутренних и мембранных 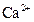 -«часов» [89].
-«часов» [89].
Автором данной диссертационной работы с помощью объединения модели кальциевой динамики с ЭК-моделью RyR-каналов впервые учтены кооперативные механизмы кинетики RyR-каналов и описан стохастический характер поведения этих каналов в процессе заполнения/высвобождения СР.
В отличие от ML-модели эффекты, связанные со стохастическим характером динамики каналов в модифицированной модели, позволяют объяснить причину увеличения амплитуды высвобождающегося Са2+ в диадное пространство с ростом частоты осцилляций изолированных внутренних Са2+-«часов», что в свою очередь позволяет на молекулярном уровне впервые описать эффект Боудича.
ЭК-модель также естественным образом вводит флуктуации частоты и амплитуды колебаний системы и шумы, нарушающие гармонический режим в модах 1 и 2. Следует отметить, что эти флуктуации заметно подавляются с ростом константы k конформационного взаимодействия между соседними RyR-каналами.
Date: 2015-08-24; view: 472; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |