Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Индексы состояния экосистем
Содержание
1 Индексы состояния экосистем. 1
Заключение. 7
Список использованных источников. 8
Индексы состояния экосистем
В связи с увеличением вычислительных мощностей, бурным развитием математического моделирования и параллельным увеличением антропогенных нагрузок на природу, возрос интерес экологов к реакциям экосистем на нарушения. А так как устойчивость и есть мера чувствительности экосистемы к нарушению, то, соответственно, необходимо выделить математически пригодный показатель, по которому можно будет ее оценивать.
Признаками неблагоприятных изменений в экосистемах могут быть: укорочение пищевых цепей, снижение уровня организации, разнообразия, повышение продуктивности, главным образом, в основании пищевой цепи. При значительных стрессовых воздействиях наблюдается однонаправленная деградация экосистемы, проявляющаяся в виде “стрессового синдрома”:
· снижение уровня первичной продуктивности (исключая случаи сильного эвтрофирования),
· уменьшение запаса биогенных веществ,
· снижение видового разнообразия,
· доминирование видов с коротким жизненным циклом,
· увеличение амплитуды колебаний численности ключевых видов,
· ретрогрессия биотической структуры (процесс, обратный сукцессии, при котором специализированные виды замещаются поливалентными формами).
На первый взгляд очевидно, что для оценки состояния экосистемы следует избрать измеряемые параметры и установить допустимые пределы их изменчивости, внутри которых систему можно считать ненарушенной. За последние десятилетия было предложено множество таких показателей, но возникло и множество проблем, связанных с их применением. Во-первых, к каждой экосистеме следует подходить индивидуально, поскольку естественная изменчивость экосистем зависит от труднообозримого числа факторов и границы изменчивости каждого параметра экосистемы также индивидуальны. Вторую проблему представляют процессы сукцессии: при изменении условий, прежняя экосистема сменяется практически новой, адаптированной к новым условиям, следовательно, показатели состояния экосистемы должны быть достаточно динамичны, чтобы быть приложимы и к новому состоянию. Наконец, практически каждый исследователь выбирает собственный показатель в зависимости от своих вкусов и опыта.
Граничные условия, которым должны соответствовать индексы состояния экосистемы, можно сформулировать следующим образом (Зилов, 2004). Они должны:
· не зависеть от состояния отдельных видов;
· иметь числовое, желательно лишенное размерности, выражение;
· быть приложимы к экосистемам разного иерархического уровня организации;
· определяться по минимальному числу наблюдений;
· иметь теоретическое обоснование.
Г. Е. Михайловский при оценке состояния планктонных сообществ предлагает измерять организованность биоценоза как
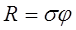 , ,
|
т. е. произведение величин сложности биоценоза и его жесткости. Сложность рассчитывается как
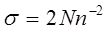 , ,
|
где N – число достоверно выявленных корреляционных связей, n – число групп планктонтов. Жесткость
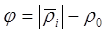 , ,
|
т. е. средний модуль достоверно выявленных корреляционных связей за вычетом порога доверительности. Очевидно, что проведение корреляционного анализа для достаточно сложного сообщества – занятие весьма трудоемкое и требующее наличия достаточно длинных рядов наблюдений.
Дж. Карром предложен Индекс Биологической Целостности (ИБЦ), рассчи-тываемый как сумма пределов изменчивости 12 характеристик сообщества рыб:
· обилие видов и их относительная численность (общее количество видов, численность чувствительных к загрязнению видов, доля стенобионтных видов);
· трофическая структура (доли всеядных, насекомоядных, хищных видов; общая численность на один отлов);
· плотность рыб;
· состояние популяций (доли гибридных форм, больных особей, особей со злокачественными новообразованиями, поврежденными плавниками, аномалиями скелета и т.п.).
На основании ИБЦ был разработан Прогностический Индекс Биологической Целостности (ПИБЦ). Он рассчитывается по регрессионным уравнениям, связывающим ИБЦ с характером донных грунтов (долевое отношение площадей песок/ил, глинистых участков), средней шириной рек, долей площади заводей и т.п. Кроме того, широкое применение нашел и Макробентосный Биотический Индекс, основанный на тех же принципах, что и ИБЦ. Все три индекса официально приняты во многих штатах США.
Г. Г. Миничевой разработан и успешно использован метод оценки и прогноза состояния растительных донных сообществ, основанный на определении удельной фотосинтетической поверхности.
К сожалению, индексы и Дж. Карра, и Г. Г. Миничевой ориентированы на оценку состояния экосистемы по состоянию одной из составляющих её подсистем: сообщества рыб, зообентоса или фитобентоса. Индексы Дж. Карра, кроме того, нарушают и первое граничное условие для показателей состояния экосистем, т.к. зависят от состояния популяций отдельных видов.
Роберт Костанца предложил показатель, названный им индексом общего здоровья системы (“overall system Health Index”, HI). Он рассчитывается как
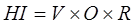 , ,
|
где V – “бодрость” экосистемы (мера активности системы, метаболизма, или первичной продукции), О – индекс организации системы, варьируищий от 0 до 1 и отражающий разнообразие и связанность системы, R – индекс эластичности системы, также варьирующий между 0 и 1.
Оценку используемых в формуле параметров Р. Костанца предлагает производить следующим образом:
· “бодрость” – прямыми измерениями валовой первичной продукции, чистой первичной продукции, продуктивности экосистемы, метаболизма;
· индекс организации системы – анализом пищевых сетей;
· эластичность системы – математическим моделированием.
Понятно, что такая оценка степени здоровья экосистемы – занятие очень трудоемкое, дорогостоящее и длительное. Кроме того, моделирование, используемое для оценки эластичности системы, вносит существенную степень неопределенности.
Исходя из концепции максимизации потока полезной энергии (в предположении, что “удачливой” системой становится та, которая сможет развить в себе способность максимизировать поток полезной энергии) вытекает введенное Г. Т. Одумом понятие энмергии или воплощенной энергии. Энмергия – это поток биогеохимической энергии через компонент, выраженный в эквивалентах солнечной энергии (другими словами эта величина количественно отражает объем энергии, потребной для создания единицы биомассы на разных трофических уровнях). Согласно законам термодинамики каждый трофический уровень способен использовать ~ 10% от энергии предшествующего трофического уровня. Наглядно это можно представить так, как это сделано в таблице 5.1.
Таблица 5.1 – Соотношение энергетических единиц (кДж м-2) для разных трофических уровней
| солнечная радиация | трофический уровень I | трофический уровень II | трофический уровень III | |
| Поток энергии | ||||
| Солнечный эквивалент | ||||
| Энмергия |
В то же время необходимо помнить о том, что поток энергии в экосистеме невозможен без вещества и информации, и чем выше уровень информации, тем выше степень использования вещества и энергии для дальнейшего удаления экосистемы от термодинамического равновесия. Максимальный поток энергии, как и предложенный на его основе показатель – энмергия, далеко не всегда отражает уровень организованности экосистемы. Кроме того, расчет энмергии достаточно сложен, т.к. её значение определяется трофическим уровнем, к которому принадлежит тот или иной организм, что требует детального анализа рационов всех компонентов экосистемы и очень трудно выполнимо для всеядных, детритоядных организмов и редуцентов.
“Информация – это то, что характеризует и отличает одну определенную структуру элементов среди огромного числа возможных структур.... Любая мера информации есть мера “уникальности” по отношению числу возможных состояний и потому она связана с энтропией ” (Маргалеф, 1992). Снижение уровня энтропии соответствует увеличению количества информации и, соответственно, уменьшение содержания информации в системе влечет за собой рост энтропии. Энтропия, или мера беспорядка, измеряется как энергия, которую нельзя получить в первоначальной форме
Любая система, и экосистема в том числе, спонтанно двигается в направлении увеличения беспорядка, т. е. энтропии, согласно второму закону термодинамики. Из этого следует, что экосистема может поддерживать свою организованность тогда и только тогда, когда она способна “сбрасывать” излишки энтропии в окружающую среду. Т.е., необходимым условием существования экосистемы является наличие источника низкоэнтропийной энергии (солнечная радиация) и “слива” высокоэнтропийной энергии (тепловое излучение). Таким образом, наиболее удачливыми системами являются те, что быстрее всех удаляются от термодинамического равновесия, то есть минимизируют энтропию.
Эксэргия определяется как расстояние между текущим состоянием системы и её состоянием в термодинамическом равновесии с окружающей средой, измеренное в единицах информации и/или энергии, т. е., эксэргия – величина отклонения системы от равновесного состояния. Она указывает на количество работы, затраченной на создание данной системы из первичных компонентов (в случае экосистемы – из молекул исходных веществ), и информации, использованной при этом, или, другими словами, на количество энергии, затрачиваемой на разрушение данной системы до первичных компонентов.
Для выявления динамики состояния экосистем служит структурная эксэргия. Структурная эксэргия – это эксэргия, отнесенная к общей биомассе. Она отражает способность экосистемы усваивать поток энергии извне, служа одновременно индикатором степени развития экосистемы, её сложности и уровня эволюционного развития организмов, из которых та состоит.
Эксэргия и структурная эксэргия рассчитываются по формулам (Зилов, 2004):
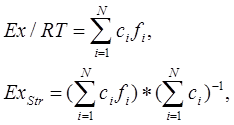
где Ех – эксэргия, R – газовая константа (8,31 Дж*моль-1*К-1), T – абсолютная температура, сi – концентрация в экосистеме компонента i, f i – пересчетный коэффициент (рассчитывается как отношение общей величины эксэргии к таковой для детрита (7,34*105)).
Соответствующие пересчетные коэффициенты уже рассчитаны для многих систематических групп организмов и опубликованы (для зеленых одноклеточных водорослей он составляет 3,4, для дрожжей – 6,4, олигохет – 35, коловраток –30, копепод – 44 и т.д.).
Если мы сравним структурную эксэргию с рассмотренными ранее параметрами состояния экосистемы, то легко увидим, что,
· во-первых, её расчет относительно прост,
· во-вторых, для его осуществления вполне достаточно данных, снимаемых в ходе обычного мониторинга,
· в-третьих, она имеет неплохое теоретическое обоснование и полностью соответствует критериям индекса состояния экосистемы, перечисленным выше.
Кроме того, основные закономерности поведения эксэргии и структурной эксэргии экосистем при внешних воздействиях были описаны в экспериментах с моделями водоемов и водотоков (моделировались эвтрофирование и токсификация). Работоспособность структурной эксэргии в качестве индикатора состояния экосистем была апробирована в лабораторных опытах (Silow, Oh, 2002), в натурных экспериментах на оз. Байкал, проведенных НИИ биологии при ИГУ (Зилов, 2004), а также с математическими моделями экосистемы оз. Байкал.
Расчет структурной эксэргии для бентосных сообществ фонового района Байкала и района сброса очищенных сточных вод БЦБК, для которого по другим показателям установлена негативность происшедших изменений, показал, что в районе сброса данный показатель существенно ниже, чем в фоновом районе.
Таким образом, представляется, что такой показатель как структурная эксэргия уже сейчас можно использовать в качестве индекса состояния экологической системы и степени его антропогенного изменения.
Естественно, он не лишен недостатков, среди которых главные – высокая степень приблизительности оценки, неспособность реагировать на замену, например, одного вида диатомей другим, или одного вида лососевых рыб другим, но он имеет такие важные преимущества как простота расчета и работоспособность.
Заключение
В работе был дан обзор индексов, используемых для оценки состояния экосистем, в котором было показано, что структурная эксэргия является одним из наиболее просто рассчитываемых и работоспособных индексов, апробированных к тому же в натурных экспериментах на оз. Байкал.
Представляется, что такой показатель как структурная эксэргия уже сейчас можно использовать в качестве оценки устойчивости экологической системы и степени ее антропогенного изменения.
| <== предыдущая | | | следующая ==> |
| Общая оценка напряженности трудового процесса | | | Введение. Оценка физического состояния |
Date: 2015-07-27; view: 689; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |