Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Показатели дыхательной функции крови
1. Кислородная емкость гемоглобина. Величина, отражающая количество кислорода, которое может связаться с гемоглобином при его полном насыщении, называется кислородной емкостью гемоглобин а.
2. Содержание кислорода в крови. Другим показателем дыхательной функции крови является содержание кислорода в крови, которое отражает истинное количество кислорода, как связанного с гемоглобином, так и физически растворенного в плазме.
3. Степень насыщения гемоглобина кислородом. В 100 мл артериальной крови в норме содержится 19-20 мл кислорода, в таком же объеме венозной крови — 13-15 мл кислорода, при этом артерио-венозная разница составляет 5-6 мл. Отношение количества кислорода, связанного с гемоглобином, к кислородной емкости последнего является показателем степени насыщения гемоглобина кислородом. Насыщение гемоглобина артериальной крови кислородом у здоровых лиц составляет 96%.
Образование оксигемоглобина в легких и его восстановление в тканях находится в зависимости от парциального напряжения кислорода крови: при его повышении. Насыщение гемоглобина кислородом возрастает, при понижении — уменьшается. Эта связь носит нелинейный характер и выражается кривой диссоциации оксигемоглобина, имеющей S-образную форму (рис.8.7).
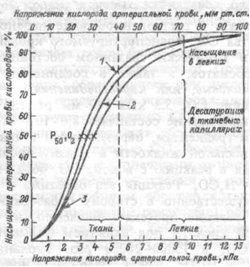 Рис.8.7. Кривая диссоциации оксигемоглобина.
Рис.8.7. Кривая диссоциации оксигемоглобина.
Рис.8.7. Кривая диссоциации оксигемоглобина.
1 — при увеличении рН, или уменьшении температуры, или уменьшении 2,3-ДФГ;
2 — нормальная кривая при рН 7,4 и 37°С;
3 — при уменьшении рН или увеличении температуры или увеличении 2,3-ДФГ.
Оксигенированной артериальной крови соответствует плато кривой диссоциации, а десатурированной крови в тканях — круто снижающаяся ее часть. Пологий подъем кривой в верхнем ее участке (зона высокого напряжения О2) свидетельствует, что достаточно полное насыщение гемоглобина артериальной крови кислородом обеспечивается даже при уменьшении напряжения О2 до 9.3 кПа (70 мм рт.ст.). Понижение напряжения О,, с 13.3 кПа на 2.0-2.7 кПа (со 100 на 15-20 мм рт.ст.) практически не отражается на насыщении гемоглобина кислородом (НЬО2 снижается при этом на 2-3%). При более низких значениях напряжения О2 оксигемоглобин диссоциирует значительно легче (зона крутого падения кривой). Так, при снижении напряжения О2 с 8.0 до 5.3 кПа (с 60 до 40 мм рт.ст.) насыщение гемоглобина кислородом уменьшается приблизительно на 15%.
Положение кривой диссоциации оксигемоглобина количественно принято выражать парциальным напряжением кислорода, при котором насыщение гемоглобина составляет 50% (Р50). Нормальная величина Р50 при температуре 37°С и рН 7.40 — около 3.53 кПа (26.5 мм рт.ст.).
Кривая диссоциации оксигемоглобина при определенных условиях может смещаться в ту или иную сторону, сохраняя S- образную форму, под влиянием изменения рН, напряжения СО2 температуры тела, содержания в эритроцитах 2,3-дяфосфоглицерата (2,3-ДФГ), от которых зависит способность гемоглобина связывать кислород. В работающих мышцах в результате интенсивного метаболизма повышается образование СО2 и молочной кислоты, а также возрастает теплопродукция. Все эти факторы понижают сродство гемоглобина к кислороду. Кривая диссоциации при этом сдвигается вправо (рис.8.7), что приводит к более легкому освобождению кислорода из оксигемоглобина, и возможность потребления тканями кислорода увеличивается. При уменьшении температуры, 2,3-ДФГ, снижении напряжения СО, и увеличении рН кривая диссоциации сдвигается влево, сродство гемоглобина к кислороду возрастает, в результате чего доставка кислорода к тканям уменьшается.
Транспорт кровью углекислого газа
Являясь конечным продуктом обмена веществ, СО2 находится в организме в растворенном и связанном состоянии. Коэффициент растворимости СО2 составляет 0.231 ммольл-1 * кПа-1 (0.0308 ммольл-1 * мм рт.ст-1.), что почти в 20 раз выше, чем у кислорода. Однако, в растворенном виде переносится меньше 10% всего количества СО,, транспортируемого кровью. В основном, СО, переносится в химически связанном состоянии, главным образом, в виде бикарбонатов, а также в соединении с белками (так называемые карбоминовые, или карбосоединения).
В артериальной крови напряжение СО2 5.3 кПа (40 мм рт.ст.), в интерстициальной жидкости его напряжение составляет 8.0- 10.7 кПа (60-80 мм рт.ст.). Благодаря этим градиентам, образующийся в тканях СО2 переходит из интерстициальной жидкости в плазму крови, а из нее — в эритроциты. Вступая в реакцию с водой, СО2 образует угольную кислоту: СО2 + Н2О <> Н2СО3. Реакция эта обратима и в тканевых капиллярах идет преимущественно в сторону образования Н2СО3 (рис.8.8.А). В плазме эта реакция протекает медленно, но в эритроцитах образование угольной кислоты под влиянием фермента ускоряет реакцию гидратации СО2 в 15000-20000 раз. Угольная кислота диссоциирует на ионы Н+ и НСО3. Когда содержание ионов НСО3 повышается, они диффундируют их эритроцита в плазму, а ионы Н+ остаются в эритроците, так как мембрана эритроцита сравнительно непроницаема для катионов. Выход ионов НСО3 в плазму уравновешивается поступлением из плазмы ионов хлора. При этом в плазме высвобождаются ионы натрия, которые связываются поступающими из эритроцита ионами НСО3, образуя NaHCO3. Гемоглобин и белки плазмы, проявляя свойства слабых кислот, образуют соли в эритроцитах с калием, а в плазме с натрием. Угольная кислота обладает более сильными кислотными свойствами, поэтому при ее взаимодействии с солями белков ион Н+ связывается с белковым анионом, а ион НСО3 с соответствующим катионом образует бикарбонат (в плазме NaHCO3, в эритроците КНСО3).
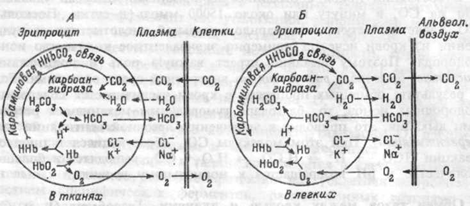 Рис.8.8. Схема процессов, происходящих в плазме и эритроцитах при газообмене в тканях (А) и легких (Б).
Рис.8.8. Схема процессов, происходящих в плазме и эритроцитах при газообмене в тканях (А) и легких (Б).
В крови тканевых капилляров одновременно с поступлением СО2 внутрь эритроцита и образованием в нем угольной кислоты происходит отдача кислорода оксигемоглобином. Восстановленный гемоглобин представляет собой более слабую кислоту (т.е. лучший акцептор протонов), чем оксигенированный. Поэтому он легче связывает водородные ионы, образующиеся при диссоциации угольной кислоты. Таким образом, присутствие восстановленного гемоглобина в венозной крови способствует связыванию СО2 тогда как образование оксигемоглобина в легочных капиллярах облегчает отдачу углекислого газа.
В переносе кровью СО2 большое значение имеет также химическая связь СО2 с конечными аминогруппами белков крови, важнейший из которых — глобин в составе гемоглобина. В результате реакции с глобином образуется так называемый карбаминогемоглобин. Восстановленый гемоглобин обладает большим сродством к СО2, чем оксигемоглобин. Таким образом, диссоциация оксигемоглобина в тканевых капиллярах облегчает связывание СО2, а в легких образование оксигемоглобина способствует выведению углекислого газа.
Из общего количества СО,, которое может быть извлечено из крови, лишь 8-10% СО, находится в соединении с гемоглобином. Однако, роль этого соединения в транспорте СО2 кровью достаточно велика. Примерно 25- 30% СО2, поглощаемого кровью в капиллярах большого круга, вступает в соединение с гемоглобином, а в легких — выводится из крови.
Когда венозная кровь поступает в капилляры легких, напряжение СО2 в плазме снижается и находящийся внутри эритроцита в физически растворенном виде СО2 выходит в плазму. По мере этого, Н2СО3 превращается в СО2 и воду (рис.8.8.Б), причем карбоангидраза катализирует реакцию, идущую в этом направлении. Н2СО3 для такой реакции доставляется в результате соединения ионов НСО3 с ионами водорода, высвобождающихся из связи с белковыми анионами.
В состоянии покоя с дыханием из организма человека удаляется 230 мл СО2 в минуту или около 15000 ммоль в сутки. Поскольку СО2 является «летучим» ангидридом угольной кислоты, при его удалении из крови исчезает примерно эквивалентное количество ионов водорода. Поэтому дыхание играет важную роль в поддержании кислотно-щелочного равновесия во внутренней среде организма. Если в результате обменных процессов в крови увеличивается содержание водородных ионов, то, благодаря гуморальным механизмам регуляции дыхания, это приводит к увеличению легочной вентиляции (гипервентиляции). При этом молекулы СО2, образующиеся в процессе реакции НСО3 + Н+ -> Н2СО3 -> Н2О + СО2, выводятся в большем количестве и рН возвращается к нормальному уровню.
Обмен газов между кровью и тканями
Газообмен О2 и СО2 между кровью капилляров большого круга и клетками тканей осуществляется путем простой диффузии. Перенос дыхательных газов (О2 — из крови в ткани, СО2 — в обратном направлении) происходит под действием концентрационного градиента этих газов между кровью в капиллярах и интерстициальной жидкостью. Разность напряжения О2 по обе стороны стенки кровеносного капилляра, обеспечивающая его диффузию из крови в интерстициальную жидкость, составляет от 30 до 80 мм рт.ст. (4.0-10.7 кПа). Напряжение СО2 в интерстициальной жидкости у стенки кровеносного капилляра на 20-40 мм рт.ст. (2.7-5.3 кПа) больше, чем в крови. Поскольку СО2 диффундирует примерно в 20 раз быстрее, чем кислород, удаление СО2 происходит гораздо легче, чем снабжение кислородом.
На газообмен в тканях влияют не только градиенты напряжения дыхательных газов между кровью и интерстициальной жидкостью, но также площадь обменной поверхности, величина диффузионного расстояния и коэффициенты диффузии тех сред, через которые осуществляется перенос газов. Диффузионный путь газов тем короче, чем больше плотность капиллярной сети. В расчете на 1 мм3 суммарная поверхность капиллярного русла достигает, например, в скелетной мышце 60 м2, а в миокарде — 100 м2. Площадь диффузии определяет также количество эритроцитов, протекающих по капиллярам в единицу времени в зависимости от распределения кровотока в микроциркуляторном русле. На выход О2 из крови в ткань влияет конвекция плазмы и интерстициальной жидкости, а также цитоплазмы в эритроцитах и клетках ткани. Диффундирующий в ткани О2потребляется клетками в процессе тканевого дыхания, поэтому разность его напряжения между кровью, интерстициальной жидкостью и клетками существует постоянно, обеспечивая диффузию в этом направлении. При увеличении потребления тканью кислорода его напряжение в крови уменьшается, что облегчает диссоциацию оксигемоглобина.
Количество кислорода, которое потребляют ткани, в процентах от общего содержания его в артериальной крови называется коэффициентом утилизации кислорода. В покое для всего организма коэффициент утилизации кислорода равен примерно 30-40%. Однако, при этом потребление кислорода в различных тканях существенно отличается, и коэффициент его утилизации, например, в миокарде, сером веществе мозга, печени, составляет 40-60%. В состоянии покоя серым веществом головного мозга (в частности, корой больших полушарий) потребляется в минуту от 0.08 до 0.1 мл О2 на 1 г ткани, а в белом веществе мозга — в 8-10 раз меньше. В корковом веществе почки среднее потребление О2 примерно в 20 раз больше, чем во внутренних участках мозгового вещества почки. При тяжелой физической нагрузке коэффициент утилизации О2 работающими скелетными мышцами и миокардом достигает 90%.
Кислород, поступающий в ткани, используется в клеточных окислительных процессах, которые протекают на субклеточном уровне с участием специфических ферментов, расположенных группами в строгой последовательности на внутренней стороне мембран митохондрий. Для нормального хода окислительных обменных процессов в клетках необходимо, чтобы напряжение О2 в области митохондрий было не меньше 0.1-1 мм рт.ст. (13.3-133.3 кПа).
Эта величина называется критическим напряжением кислорода в митохондриях. Поскольку единственных резервом О2 в большинстве тканей служит его физически растворенная фракция, снижение поступления О2 из крови приводит к тому, что потребности тканей в О2 перестают удовлетворяться, развивается кислородное голодание и окислительные обменные процессы замедляются.
Единственной тканью, в которой имеется депо О2, является мышечная. Роль депо О2 в мышечной ткани играет пигмент миоглобин, близкий по строению к гемоглобину и способный обратимо связывать О2. Однако, содержание миоглобина в мышцах человека невелико, и поэтому количество запасенного О, не может обеспечить их нормальное функционирование в течение длительного промежутка времени. Сродство миоглобина к кислороду выше, чем у гемоглобина: уже при напряжении О, 3-4 мм рт.ст. 50% миоглобина переходит в оксимиоглобин, а при 40 мм рт.ст. миоглобин насыщен О2 до 95%. Во время сокращения мышцы, с одной стороны, увеличиваются потребности клеток в энергии и усиливаются окислительные процессы, с другой — резко ухудшаются условия доставки кислорода, поскольку при сокращении мышца сдавливает капилляры и доступ крови по ним может прекращаться. Во время сокращения расходуется О2, запасенный в миоглобине за время расслабления мышцы. Особое значение это имеет для постоянно активно работающей мышцы сердца, поскольку ее снабжение кислородом из крови носит периодический характер. Во время систолы в результате повышения интрамурального давления кровоток в бассейне левой коронарной артерии снижается и во внутренних слоях миокарда левого желудочка может на короткое время полностью прекратиться. Когда напряжение О2 в мышечных клетках падает ниже 10-15 мм рт.ст. (1.3-2.0 кПа), миоглобин начинает отдавать О,, запасенный в виде оксимиоглобина за время диастолы. Среднее содержание мио глобина в сердце составляет 4 мг/г. Поскольку 1 г миоглобина может связать примерно до 1.34 мл кислорода, в физиологических условиях запасы кислорода в миокарде составляют около 0.005 мл на 1 г ткани. Этого количества кислорода достаточного для того, чтобы в условиях полного прекращения его доставки кровью поддерживать в миокарде окислительные процессы лишь в течение 3-4 с. Однако, длительность систолы намного короче, поэтому миоглобин, выполняющий функцию кратковременного депо О2, предохраняет миокард от кислородного голодания.
Date: 2015-07-27; view: 3199; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |