Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Механизм
Современная механистическая теория морфогенеза приписывает главную роль ДНК по следующим четырем причинам. Во-первых, было обнаружено, что многие случаи наследственных различий между животными или растениями данного вида зависят от генов, которые действительно могут быть «картированы», и может быть установлено их местонахождение в определенных участках определенных хромосом. Во-вторых, известно, что химической основой генов является ДНК и что их специфичность зависит от последовательности пуриновых и пиримидиновых оснований в ДНК. В-третьих, ясно, каким образом может действовать ДНК как химическая основа наследственности: с одной стороны, она служит шаблоном для своей собственной репликации[48] благодаря специфичности связывания в пары оснований в ее двух комплементарных нитях; с другой стороны, она служит шаблоном для последовательности аминокислот в белках. Последнюю роль она играет не прямо, а через посредника; одна из ее нитей вначале «транскрибируется» (переписывается.— Прим. пер.), образуя одну нить молекулы «передаточной» РНК (так называемой РНК-мессенджера), с которой в процессе синтеза белка «считывается» последовательность оснований по три сразу. Различные триплеты оснований придают специфические свойства различным аминокислотам, и, таким образом, генетический код «переводится» в последовательность аминокислот, которые соединяются с образованием характерных полипептидных цепей; они, в свою очередь, сворачиваются с образованием белков. Наконец, характеристики клетки зависят от ее белков: метаболизм и способность к химическому синтезу белков, некоторые из ее структур — от структурных белков, а свойства поверхности, которые позволяют ее «узнавать» другим клеткам,— от специальных поверхностных белков.
В рамках механистического образа мышления центральной проблемой развития и морфогенеза считается контроль синтеза белка. В бактериях специфические вещества, называемые стимуляторами (индукторами), могут вызывать транскрипцию специфических участков ДНК в РНК-мессенджере, которая затем служит шаблоном для синтеза специфических белков. Классическим примером является индукция фермента галактозидазы лактозой в бактерии Escherichia coli. «Включение» гена происходит через сложную систему, содержащую репрессорный белок, который блокирует транскрипцию путем связывания со специфическим участком ДНК; его способность к такой блокировке существенно уменьшается в присутствии химического индуктора. Подобным же образом осуществляется «выключение» гена специфическими химическими репрессорами. У животных и растений система «включения» и «выключения» генов более сложная и сейчас еще не вполне понятная. Дополнительные трудности возникают вследствие недавно установленного факта, что РНК-мессенджер может быть составлена из частей, транскрибированных (переписанных) с различных участков ДНК и затем специфическим образом соединенных вместе. Более того, синтез белков также контролируется на «трансляционном уровне»; синтез белка может «включаться» и «выключаться» различными факторами даже в присутствии подходящей РНК-мессенджере.
Таким образом, разные белки, производимые различными типами клеток, зависят от способа контроля синтеза белка. Единственный способ объяснения этого с механистических позиций — через физико-химические воздействия на клетки; следовательно, модели дифференциации должны зависеть от физико-химических связей в ткани. Природа этих воздействий неизвестна, и выдвигались разные варианты: градиенты концентраций специфических химических реагентов, диффузионно-реакционные системы с химической обратной связью; электрические градиенты; электрические или химические колебания (осцилляции); механические контакты между клетками или другие факторы или же комбинации различных факторов. При этом клетки должны реагировать на эти различия характерным образом. Один из существующих сейчас способов рассмотрения этой проблемы состоит в том, чтобы считать эти физические или химические факторы источниками «позиционной информации», которую клетки затем «интерпретируют» в соответствии со своей генетической программой путем «включения» синтеза определенных белков[49].
Эти различные аспекты центральной проблемы контроля синтеза белка интенсивно исследуются в настоящее время. Большинство механистически мыслящих биологов надеются, что решение этой проблемы приведет или по крайней мере продвинет нас к объяснению морфогенеза с чисто механистических позиций.
 Чтобы оценить, насколько такое механистическое объяснение морфогенеза вероятно или даже возможно, следует рассмотреть одно за другим несколько препятствий:
Чтобы оценить, насколько такое механистическое объяснение морфогенеза вероятно или даже возможно, следует рассмотреть одно за другим несколько препятствий:
(1) Возможности объяснения, приписываемые ДНК и синтезу специфических белков, существенно ограничены тем фактом, что как ДНК, так и белки различных типов могут быть очень похожи. Например, при детальном сопоставлении белков человека и шимпанзе значительное число белков оказались идентичными а другие имели лишь незначительные различия: «Определение последовательности (сиквенс) аминокислот, иммунологические методы и электрофорез дают согласующиеся оценки генетического подобия. Все эти подходы указывают на то, что средний полипептид человека более чем на 99 процентов идентичен своему обезьяньему двойнику»[50]. Сравнения так называемых неповторяющихся последовательностей ДНК (то есть частей, которые считаются генетически значимыми) показывают, что суммарное различие в последовательностях ДНК человека и шимпанзе составляет всего лишь 1,1 процента.
Подобные же сравнения различных видов мышей или плодовых мушек дрозофил обнаружили более значительные различия между этими близко связанными видами, чем между людьми и шимпанзе, что приводит к следующему выводу: «Контрасты между эволюцией молекул и организмов показывают, что эти два процесса в большой степени независимы друг от друга»[51].
Допустим, однако, в порядке дискуссии, что наследственные различия между видами столь различными, как человек и шимпанзе, действительно можно объяснить, предположив очень малые изменения в структуре белков, или наличие различных белков в малых количествах, или генетические изменения, которые влияют на контроль синтеза белка (возможно, зависящий до некоторой степени от различий в расположении ДНК в хромосоме), или через комбинации этих факторов.
В одном организме развитие происходит в разных формах, хотя ДНК одна и та же. Рассмотрим, например, руку и ногу человека: обе содержат клетки одинакового вида (клетки мускулов, клетки соединительной ткани и т. д.) с идентичными белками и идентичной ДНК. Таким образом, различия в форме руки и ноги не могут быть приписаны ДНК как таковой; они должны быть приписаны факторам, определяющим строение органа, которые действуют различно в развивающейся руке или ноге. Высокая степень организованности в структуре тканей — например, соединение сухожилий с правой стороной кости — показывает, что эти определяющие строение факторы должны действовать с высокой точностью. Механистическая теория жизни полагает, что эти факторы должны быть физико-химическими по природе. Однако на сегодняшний день их природа неизвестна.
Даже если физические или химические факторы, определяющие процесс дифференциации, могут быть идентифицированы, все же остается проблема выяснения того, каким образом сами эти факторы организованы в начальный момент. Эта проблема может быть проиллюстрирована на двух из очень немногочисленных случаев, в которых химические «морфогены» действительно были выделены.
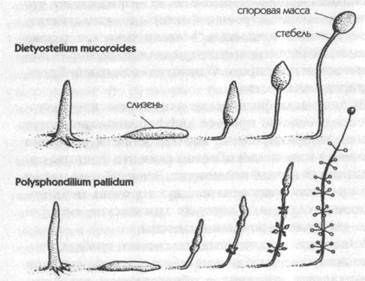
Рис. 4. Миграционная и кульминационная стадии двух видов плесневых грибков. Слева показаны вновь развившиеся сложные организмы, образованные агрегацией множества свободно живущих амебоидных клеток. Они мигрируют как «слизни» и затем растут вверх, дифференцируясь в стебли, несущие тела спор (Bonner, 1958)
Во-первых, в клеточных плесневых грибках свободно живущие амебоидные клетки агрегируют при определенных условиях с образованием «слизня», который после движения по земле в течение некоторого времени растет вверх, в воздух, и дифференцируется в стебель, несущий споровую массу (рис. 4). Было показано, что агрегация этих клеток зависит от относительно простого химического реагента, АМФ (аденозин 3', 5'-монофосфата). Но в образовавшемся составном организме, хотя распределение циклического АМФ связано с картиной дифференциации, «неясно, является ли распределение АМФ причиной или следствием предшествующей дифференциации». Более того, даже если это вещество играет ключевую роль в дифференциации, оно само не может объяснить ни того, почему оно распределено именно таким образом, ни того факта, что это распределение изменяется от одного вида к другому: какие-то другие факторы должны быть ответственны за сложную картину его распределения. Существует большое разнообразие мнений относительно возможной природы этих факторов[52].
Во-вторых, известно, что в высших растениях гормон ауксин (индолил-3-ускусная кислота) играет роль в контроле дифференциации сосудов. Но тогда что определяет производство и распределение ауксина? Возможный ответ может быть: сама дифференциация сосудов. По-видимому, ауксин выделяется дифференцирующимися сосудистыми клетками как побочный продукт расщепления белков, которое происходит по мере взросления (развития) клеток. Таким образом, система замкнута: она помогает сохранять картины дифференциации, но не объясняет, как они возникли[53].
Допустим, однако, в порядке дискуссии, что было бы возможно идентифицировать, какие факторы создают картину физических или химических воздействий, которые, в свою очередь, определяют картину дифференциации; предположим также, что можно определить и те способы, с помощью которых контролируются сами эти контролирующие факторы, и так далее. Тогда возникает проблема регуляции: если часть системы удалена, эта сложная последовательность физико-химических связей должна быть нарушена. Но оказывается, что каким-то образом оставшиеся части системы могут изменить свой обычный путь развития и развиваться дальше с достижением более или менее нормального конечного результата.
Обычно все согласны в том, что это проблема чрезвычайно трудная; она еще далеко не понята даже в общих чертах. Защитники механистической теории надеются, что она может быть решена с помощью большой, кропотливой работы; их оппоненты отрицают, что она может быть даже в принципе решена механистическим путем. Однако обсуждения ради можно еще раз предположить, что механистическое решение может быть найдено.
Тогда остается другая проблема — каким образом «информация о положении» вызывает свои эффекты. Простейшая возможность состоит в том, что эта информация определяется градиентом концентрации специфического (химического) реагента и что клетки под воздействием концентрации выше некоторого значения синтезируют один набор белков, а при концентрации ниже этого порога — другой. Опять-таки допустим, что этот или другие механизмы, с помощью которых эта «позиционная информация» может быть «интерпретирована», действительно могут быть выяснены[54]. Теперь, в конце этой цепи весьма оптимистических предположений, достигается ситуация, в которой разные клетки, организованные в нужном порядке, производят различные белки.
До сих пор рассматривался набор отношений один к одному: ген «включается» специфическим стимулом, ДНК транскрибируется в РНК, а РНК переводится в определенную последовательность аминокислот, в полипептидную цепь. Но здесь простая причинная цепочка приходит к концу. Каким образом полипептидные цепи свертываются в специфические трехмерные структуры белков? Как белки приводят к появлению в клетках их характерной структуры? Как клетки агрегируют с образованием тканей с характерной структурой? И так далее. Это и есть проблемы собственно морфогенеза: синтез специфических полипептидных цепей обеспечивает основу для аппарата метаболизма и структурные материалы, от которых зависит морфогенез; но что фактически определяет организацию и структуры, в которые объединяются клетки и ткани? Механистическое толкование здесь таково, что все это может быть объяснено через физические взаимодействия и что этот процесс протекает спонтанно (самопроизвольно), при условии что нужные белки находятся в нужных местах в нужное время и в нужной последовательности. На этой ключевой стадии механистическая биология явно слагает с себя полномочия и решение проблемы морфогенеза просто возлагается на физику.
Действительно, полипептидные цепи самопроизвольно свертываются, если имеются подходящие условия, в белки с характерной трехмерной структурой. Их даже можно заставить развернуться, а затем, изменив условия, снова свернуться в пробирках, так что этот процесс не зависит от какого-либо таинственного свойства живой клетки. Более того, белковые субъединицы могут агрегировать в пробирке с образованием структур, которые в норме образуются в живых клетках: например, субъединицы белка тубулина объединяются в длинные палочкообразные структуры, называемые микротрубочками[55].
А еще более сложные структуры, такие как рибосомы, могут образоваться в результате самопроизвольной агрегации различных белков и компонентов РНК. Вещества других классов, например липиды клеточных мембран, также способны спонтанно агрегировать в пробирке.
По своей способности к самопроизвольной агрегации эти структуры напоминают кристаллы; многие из них действительно могут рассматриваться как кристаллические или квазикристаллические. Так что в принципе они представляют не большую или не меньшую проблему, чем обычная кристаллизация; можно допустить, что здесь протекают те же физические процессы.
Тем не менее все процессы морфогенеза, безусловно, нельзя рассматривать как процессы кристаллизации. Они должны включать множество других физических факторов: например, на формы, принимаемые мембранами, должны влиять силы поверхностного натяжения, а на структуры гелей и золей — коллоидные свойства их составляющих. И кроме того, некоторые формы могут возникать в результате статистически случайных флуктуации; простые примеры появления «порядка через флуктуации» начали изучать с точки зрения необратимой или неравновесной термодинамики в неорганических системах, и близкие по характеру процессы вполне могут протекать в клетках и тканях[56]. Однако механистическая теория не просто предполагает, что эти и другие физические процессы играют роль в морфогенезе; она утверждает, что морфогенез можно полностью объяснить на языке физики. Что это означает? Если что-либо наблюдаемое определяется как в принципе объяснимое физически просто потому, что оно происходит, то это должно быть так по определению. Но это необязательно означает, что оно может быть объяснено с помощью известных законов физики. В отношении биологического морфогенеза можно сказать, что это объяснение может быть достигнуто, если биолог, который знает полную последовательность оснований в ДНК организма и имеет подробное описание физико-химического состояния оплодотворенного яйца и окружающей среды, в которой оно развивалось, может предсказать, основываясь на фундаментальных законах физики (то есть квантовой теории поля, уравнениях электромагнетизма, втором законе термодинамики и т. д.), во-первых, трехмерную структуру всех белков, которые будет производить этот организм; во-вторых, ферментативные и другие свойства этих белков; в-третьих, полную картину метаболизма всего организма; в-четвертых, природу и последствия всех типов позиционной информации, которая появилась бы в процессе его развития; в-пятых, структуру его клеток, тканей и органов и форму целого организма; и наконец, для животного — его инстинктивное поведение. Если все эти предсказания могут быть успешными и если, кроме того, ход процессов регуляции и регенерации также может быть предсказан a priori, это действительно стало бы убедительной демонстрацией того, что живые организмы полностью объяснимы с помощью известных законов физики. Но, конечно, ничего подобного сегодня сделано быть не может. И нет способа продемонстрировать, что такое объяснение возможно. Его вообще может не быть.
Таким образом, если механистическая теория утверждает, что все явления морфогенеза в принципе объяснимы с помощью известных законов физики, она вполне может ошибаться: так мало понятно сейчас, что, по-видимому, нет серьезных оснований для твердого убеждения в том, что с помощью известных законов можно объяснить все явления. Но это, по крайней мере, проверяемая теория: она может быть отвергнута в результате открытия нового закона физики. С другой стороны, если бы механистическая теория утверждала, что живые организмы подчиняются как известным, так и неизвестным законам природы, тогда она была бы неопровержима; это было бы просто общим утверждением о том, что объяснение возможно. Такая теория не противостояла бы органицизму и витализму, но включала бы их.
На практике механистическая теория жизни не рассматривается как строго определенная, опровергаемая научная теория; скорее, она служит для оправдания консервативного метода работы в рамках установившегося строя мысли, предлагаемого современной физикой и химией. Хотя обычно считается, что она утверждает то, что живые организмы в принципе объяснимы через известные законы физики, если бы были открыты новые законы физики и, таким образом, они стали бы известны, механистическая теория легко могла бы быть изменена так, чтобы включить и их.
Можно ли было бы такую модифицированную теорию называть механистической или нет, это лишь вопрос определения.
Когда так мало поняты явления морфогенеза и поведения, безусловно, нельзя исключить возможность, что по крайней мере некоторые из этих явлений зависят от причинного фактора, пока еще не признанного физикой. В механистическом подходе этот вопрос просто отложен. Тем не менее он остается полностью открытым.
Date: 2015-07-25; view: 386; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |