Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Схема 8. Механизм глюконеогенеза (цикл Кори)
1 – пируваткарбоксилаза; 2 – фосфоенолпируваткарбоксилаза; 3 – фруктозо-1,6-дифосфатаза; 4 – глюкозо- 6-фосфатаза.
Важную роль в процессах глюконеогенеза играет так называемый цикл аланина, который происходит в мышечной ткани. При голодании вследствие катаболизма белков высвобождаются аминокислоты, более 50% количества которых составляет аланин. Поступая в печень, он используется там не для синтеза белка, а для образования Г-6-Ф через стадию пирувата. Некоторое количество аланина также через стадию пирувата превращается в Г-6-Ф непосредственно в мышечной ткани. В мышцах аланин образуется из пировиноградной кислоты. Основными донаторами группы NH(2) при этом являются такие аминокислоты, как лейцин, изолейцин, валин.
Таким образом, количество глюкозы в крови, оттекающей от печени, зависит в основном от двух взаимосвязанных процессов: гликолиза и глюконеогенеза, которые в свою очередь регулируются ключевыми ферментами фосфофруктокиназой и фруктозо-1, 6-бисфосфатазой соответственно. Активность этих ферментов адаптирована к пищевому и гормональному состоянию организма. Как отмечалось выше, фосфофруктокиназа является ключевым ферментом гликолиза и ее активность контролируется рядом метаболитов. За последние 10 лет многочисленными исследованиями показано, что основным таким регулятором является фруктозо-2, 6-бисфосфат – активатор фосфофруктокиназы. Прием глюкозы увеличивает количество в печени фруктозо-2, 6-бисфосфата, а глюкагон снижает его количество. Фруктозо-2, 6-бисфосфат синтезируется из фруктозо-6-фосфата и АТФ под воздействием 6-фосфофрукто-2-киназы. Этот же белок обладает также фруктозо-2, 6-бисфосфатазной активностью, которая расщепляет фруктозо-2, 6-бисфосфат на фруктозо-6-фосфат и неорганический фосфор. Таким образом, этот фермент является как бы многофункциональным и его фосфорилирование цАМФ-зависимой протеинкиназой снижает киназную и увеличивает фосфатазную активности, что приводит к уменьшению фруктозо-2, 6-бисфосфата.
Аффинность печеночной фосфофруктокиназы к фруктозо-2, 6-фосфату в 100 раз выше, чем к фруктозо-1, 6-бисфосфату и более чем в 1000 раз выше, чем к глюкозо-1, 6-бисфосфату. Помимо активирования фосфофруктокиназы фруктозо-2, 6-бисфосфат угнетает фруктозо-1, 6-бисфосфатазу, а уменьшение его количества снижает конверсию фруктозо-6-фосфата в фруктозо-1, 6-бисфосфат и повышает конверсию фруктозо-1, 6-фосфата в фруктозо-6-фосфат, что усиливает процесс глюконеогенеза.
Таким образом, метаболизм глюкозы и контроль синтеза и распада гликогена отражает баланс между инсулином и глюкагоном на определенный отрезок времени. Глюкагон, активируя цАМФ-зависимую протеинкиназу, способствует фосфорилированию фосфорилазы киназы, фосфорилазы и гликогенсинтазы, которые увеличивают процессы глюконеогенеза. Одновременное угнетение гликолиза, наблюдаемое при этом, является следствием активирования пируваткиназы, фруктозо-1, 6-бисфосфатазы и 6-фосфофрукто-2-киназы/фруктозо-2, 6-бисфосфатазы, что уменьшает количество фруктозо-2, 6-бисфосфата с ингибированием гликолиза и активированием глюконеогенеза. Инсулин же, наоборот, способствует дефосфорилированию этих ферментов, что активирует синтез гликогена и гликолиз.
Источниками ресинтеза глюкозы также служат глицерин, образующийся при обмене липидов, и некоторые промежуточные продукты цикла Кребса – лимонная, кетоглутаровая, но в большей степени щавелевоуксусная кислоты.
Обмен глюкозы частично осуществляется и через образование глюкуроната (глюкуронатный цикл), который посредством промежуточных стадий ксилулозо-5-фосфата взаимосвязан с гексозомонофосфатным циклом. Глюкуронатный цикл у всех млекопитающих, кроме человека и морской свинки, является местом эндогенного образования аскорбиновой кислоты (витамин С). Глюкуронаты образуются в толстой кишке в процессе бактериального расщепления полисахаридов, поступают в печень, где и включаются в глюкуронатный цикл.
Необходимо отметить, что частично обмен глюкозы в организме осуществляется через образование полиолов (полиоловый путь). Последние относятся к спиртам и образуются посредством отщепления карбоксильной группы альдоз и кетоз при участии альдозоредуктазы либо полиолдегидрогеназы. Этим путем глицеральдегид превращается в глицерин, глюкоза – в сорбитол, галактоза – в галактитол (дульцитол), ксилоза – в ксилит, арабиноза – в арабитол. В качестве кофермента при образовании полиолов используется NADPH. Этот обмен глюкозы происходит, в частности, в периферических нервах, где концентрация сорбитола выше, чем в других органах. Затем сорбитол при участии фермента сорбитолгидрогеназы может окисляться с образованием фруктозы. В хрусталике глаза глюкоза может непосредственно преобразоваться во фруктозу. Оба соединения (сорбитол и фруктоза) способствуют накоплению воды в тканях вследствие гиперосмолярности, что имеет патогенетическое значение в развитии поздних осложнений сахарного диабета.
Наряду с глюкозой большое значение в обеспечении организма энергией имеют жиры. При голодании энергетические расходы в основном покрываются за счет жиров, тогда как глюкоза сохраняется для снабжения энергией мозга. Жирные кислоты угнетают поглощение глюкозы мышцами. При гипогликемии происходит мобилизация жирных кислот и увеличение окисления их в мышцах при одновременном снижении утилизации глюкозы, а прием углеводов и повышение уровня глюкозы в крови приводят к снижению липолиза и усилению липогенеза. Цикл глюкоза – свободные жирные кислоты (цикл Рэндла) является одним из механизмов, обеспечивающих гомеостаз глюкозы. Концентрация кетоновых тел также имеет прямое отношение к регуляции содержания глюкозы в крови.
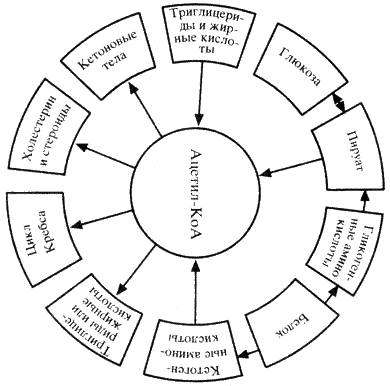
Ацетил-КоА, являясь конечным продуктом гликолитического цикла, может использоваться как источник энергии (в цикле Кребса), а также участвовать в синтезе триглицеридов, холестерина, стероидов и образовании кетоновых тел (схема 9).
Date: 2015-07-01; view: 507; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |