Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Возбудительно-тормозные отношения в спинном мозгу 3 page
Зрительная сенсорная система на таламическом уровне представлена латеральными коленчатыми телами. Они являются наиболее сложно организованными из специфических сенсорных ядер таламуса и
состоят из дорсальных и вентральных отделов. Основным релейным ядром зрительной системы является дорсальный отдел, получающий 2/3 волокон зрительного тракта; он организован в виде пластин (слоев). У человека и приматов 6 пластин, из них 1-я, 4-я и 6-я получают афферентацию от сетчатки контрала-терального глаза, а 2-я, 3-я и 5-я — от сетчатки ипси-латерального. Разные афферентные системы сетчатки раздельно проецируются в определенные пластины. Дорсальный отдел состоит из релейных тала-мокортикальных пучковидных и овоидных нейронов, аксоны которых направляются в 17-е и 18-е поля зрительной коры, и интернейронов, как тормозящих, так и возбуждающих. Рецептивные поля сетчатки, представленные в таламусе у хищников и приматов, имеют концентрическую форму и состоят из центральной зоны и окружающей ее в виде кольца периферической зоны. Одни нейроны возбуждаются при освещении центра и затемнении периферии, другие активируются освещением периферии и тормозятся освещением центра рецептивного поля. Между типами рецептивных полей нейронов наружного коленчатого тела и проецирующимися на них ганглиозными клетками сетчатки имеется полное соответствие: они мо-носинаптически избирательно связаны. Проекция центральной ямки сетчатки представлена наиболее широко и локализована в верхних слоях, проекция периферии сетчатки --в нижних слоях. Нейроны наружного коленчатого тела наряду с релейной выполняют интегративную функцию: здесь происходит сложное взаимодействие нейронов как с моно-, так и с полирецептивными полями. Вентральный отдел наружных коленчатых тел содержит нейроны с разным характером и размерами рецептивных полей: правильной концентрической формы диаметром до 60°, мел-
кие поля до 3е, обширные, захватывающие почти все контралатеральное поле зрения, цветоизбирательные поля неправильной формы.
К специфическим ядрам таламуса наряду с сенсорными проекционными ядрами относят релейные моторные ядра и ядра передней группы, составляющие единый комплекс. В моторных релейных ядрах переключается афферентация, идущая от ядер мозжечка и бледного шара к моторной коре.'Нейроны релейных моторных ядер в зависимости от афферентации пространственно организованы: в оральном отделе -от бледного шара, в каудальном — от зубчатых ядер мозжечка, в медиокаудальном - - от вестибулярной системы и мышечных веретен. Релейная функция ядер передней группы состоит в переключении импульсов из мамиллярных тел гипоталамуса в лимбическую кору; ведущая афферентная система передних ядер представлена мамиллоталамическим трактом. Передняя группа ядер входит в лимбическую систему — круг Пейпеца, включающий гиппокамп, мамилляр-ные тела, передние ядра таламуса и лимбическую кору.
5.3.1.3. Ассоциативные ядра таламуса
Ассоциативные ядра таламуса наиболее поздно дифференцируются и интенсивно развиваются в ходе эволюции зрительного бугра. Они характеризуются следующими признаками.
1. Эфферентные волокна направляются к ассоциа
тивным областям коры, а также частично и к специ
фическим проекционным областям.
2. Связь с ассоциативными областями коры моно-
синаптическая; волокна идут к I и II слоям коры, от
давая по пути коллатерали в IV и V слои и образуя
аксоматические контакты с пирамидными нейронами.
3. Основная афферентация поступает, главным об
разом, не из периферических отделов сенсорных сис
тем, а от специфических и других таламических ядер.
4. Основную массу клеток составляют нейроны 3
типов: а) мультиполярные клетки с 4—6 дендрита-
ми, имеющие тело округлой формы с наибольшим
диаметром 170 мкм (составляют около 40% всех ней
ронов ассоциативных ядер); б) биполярные клетки
веретенообразной формы (d — 10-15 мкм) с тонкими
аксонами и ветвящимися дендритами, покрытыми
шипиками — специфическими по форме синапсами;
в) трехотростчатые нейроны с коротким аксоном и
дендритами, покрытыми шипиками, которые состав
ляют свыше 20% всех клеток.
5. Электрическое раздражение ассоциативных ядер
вызывает в ассоциативных областях коры вторичные
ассоциативные положительно-отрицательные ответы,
которые не исчезают ни после разрушения первичных
корковых зон, ни после разрушения ретикулярной
формации мозгового ствола. Они также не определя
ются возбуждением, идущим в кору от неспецифи
ческих ядер таламуса.
К ассоциативным ядрам относятся: медиодорсаль-ное, латеральное дорсальное и подушка. В последнее время в их число входит также и задняя группа ядер. Важной особенностью нейронов ассоциативных ядер является способность многих из них реагировать на раздражения разных сенсорных входов, большая часть нейронов является двух- или полисенсорными. Среди нейронов ассоциативных ядер таламуса есть такие, которые реагируют только на комплексные раздражения. В медиодорсальном ядре преобладают реакции нейронов на соматическую афферентацию, в подушке — на зрительную, а в задней группе — на слуховую. Ряд нейронов подушки реагирует на движение
предмета в поле зрения и способен различать направления движения. Наличие на нейронах ассоциативных ядер полисенсорной конвергенции способствует процессам взаимодействия в них возбуждений разных сенсорностей. Это обеспечивает большие возможности для взаимодействия и интеграции в нейронах ассоциативных ядер импульсации от всех сенсорных систем. Обширные связи ассоциативных ядер таламуса с сенсорными системами и двусторонние связи с ассоциативными областями коры обеспечивают более высокий уровень интегративных процессов, чем в специфических ядрах.
5.3.1.4. Неспецифические ядра таламуса
Неспецифические ядра таламуса не относятся к какой-либо одной специфической проекционной или ассоциативной системе: они морфологически и функционально связаны со многими системами и участвуют вместе с ретикулярной формацией мозгового ствола в осуществлении неспецифических функций. Неспецифические ядра таламуса характеризуются следующими особенностями.
1. Проецируются на кору больших полушарий бо
лее диффузно, чем специфические ядра.
2. Связь с корой в основном полисинаптическая;
волокна направляются к нейронам всех слоев коры и
образуют аксодендритические синапсы на пирамид
ных нейронах.
3. Афферентация к ним поступает, главным обра
зом, из ретикулярной формации мозгового ствола, а
также гипоталамуса, лимбической системы, базаль-
ных ядер, специфических ядер таламуса, имеются и
отдельные коллатерали от сенсорных путей.
4. Нейронные сети имеют преимущественно «ре
тикулярное» строение, т.е. состоят, в основном, из
густой сети нейронов с длинными, слабо ветвящимися дендритами.
5. Одиночное электрическое раздражение, как правило, не вызывает одиночных электрических ответов в коре, ритмическая низкочастотная стимуляция вызывает (Демпси и Моррисон) реакцию вовлечения (синхронизации), а высокочастотная — реакцию активации (десинхронизации электрической активности коры). Эта реакция регистрируется из неспецифических зон коры, так как в специфических зонах она подавляется специфической импульсацией из сенсорных зон.
Эти две таламокортикальные системы находятся в постоянном взаимодействии, причем если неспецифическая система усиливает специфическую, то специфическая, наоборот, подавляет неспецифическую. Под влиянием неспецифических импульсов ответная реакция корковых нейронов на специфическую стимуляцию заметно усиливается, т.е. неспецифические таламические импульсы облегчают деятельность корковых нейронов, повышая их возбудимость. При обратном же сочетании раздражений, т.е. когда за стимуляцией специфических следует раздражение неспецифических ядер, корковые неспецифические ответы угнетаются. Этим и обусловливается сравнительно плохая проявляемость реакции вовлечения в проекционных зонах коры.
К неспецифическим ядрам таламуса относят: срединный центр, парацентральное ядро, центральное медиальное, центральное латеральное, субмедиалъ-ное, вентральное переднее, парафасцикулярный комплекс, ретикулярное, перивентикулярное и центральную серую массу. Деятельность неспецифических ядер таламуса приводит к генерации «веретенообразной* электрической активности в таламусе и
коре. Она возникает при низкочастотной стимуляции неспецифических ядер с большим латентным периодом и проявляется диффузно в виде электрических ответов в ритме стимуляции, амплитуда которых периодически изменяется, образуя веретена. Функциональное «выключение» или разрушение медиальных, интраламинарных структур и ретикулярного ядра нарушали «веретенообразную» активность. Показано, что в неспецифических ядрах имеются группировки нейронов, различающиеся своим вкладом в генерацию «веретенообразной» активности. Неспецифические ядра таламуса оказывают на кору головного мозга модулирующее влияние, регулируют ее функциональное состояние. Афферента-ция из неспецифических ядер не ведет к возникновению возбуждения в нейронах коры, а изменяет их возбудимость к специфической афферентаЦии. Деятельность неспецифических ядер таламуса тесно связана с механизмами развития сна, саморегуляции функционального состояния, интегративными процессами мозга и высшей нервной деятельности. На важную роль неспецифических ядер в этих процессах указывает и множество клинических наблюдений. Патологические процессы в этих структурах у человека или их повреждения во время операций приводили к нарушениям сознания. Различные изменения сознания получены и при электрических воздействиях на неспецифические структуры таламуса через электроды с лечебной целью. Эти факты привели к представлению о локализации в неспецифических ядрах таламуса «центроэнцефалической» системы мозга. Более подробно об участии неспецифических ядер в регуляции работы мозга будет сказано в главе о саморегуляции функционального состояния головного1 мозга.
5.3.1.5. Организация интегративных прог^ессов
В нейронных сетях и специфических, и неспецифических ядер таламуса происходят сложные интег-ративные процессы, связанные с переработкой информации, организацией взаимодействия и регуляцией. Одним из механизмов такой интеграции являются тормозные процессы: в таламических нейронах зарегистрированы длительные (около 100 мс) тормозные постсинаптические потенциалы. При участии тормозных интернейронов осуществляются процессы афферентного и возвратного торможений в нейронных сетях таламуса (рис. 5.3). После возбуждения эфферентными сигналами релейных нейронов в них по механизму возвратного торможения возникают положительные ТПСП, после которых вновь возникают возбудительные постсинаптические потенциалы. С периодическим чередованием возбуждения и возвратного торможения в релейных нейронах таламуса связана генерация ритмической электрической активности мозга, в частности, альфа- и сигма-ритма. Торможение способствует созданию пространственного кон-
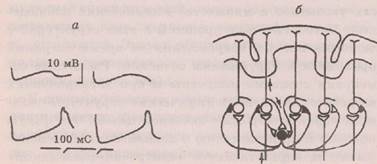
Рис, 5.3. Постсинапгпическое. торможение в нейронах тала муса: а — примеры ТПСП в релейных нейронах таламуса кошки; б —- схема возвратного торможения как механизма их возникновения (Костюк, 1977)
траста вокруг основного возбужденного очага, организует во времени реакции нейронов, обеспечивает синхронизацию нейронной активности за счет одновременного влияния на возбудимость очень многих нейронов со стороны тормозных клеток. Кроме передачи проекционных влияний на кору, нейронные ядра таламуса могут сами осуществлять замыкание рефлекторных путей без участия коры головного мозга и организацию ряда сложных рефлекторных функций. Данные о надсегментарных функциях таламуса получены на животных с полностью удаленными корой и подкорковыми ядрами, но сохраненным таламусом. У таких «таламических» животных сохраняются не только простые движения, но и сложные цепи двигательных реакций, обеспечивающих передвижение в пространстве (локомоции), сложные двигательные реакции, для которых необходима интеграция соматических мышц и мускулатуры внутренних органов (глотание, сосание, жевание). При повреждении таламуса все эти двигательные реакции нарушаются. К числу надсегментарных функций таламуса относится анализ болевой чувствительности и организация болевых реакций. Таламус является высшим центром болевой чувствительности: импульсы, идущие к нейронам таламуса от поврежденных участков тела и внутренних органов, вызывают активацию таламических нейронов и субъективные болевые ощущения. У «таламических» животных сильные раздражения сенсорных входов вызывают крик, вегетативные и поведенческие реакции. Ощущение боли связано с возбуждением нейронов неспецифических ядер таламуса; для него не обязательно участие нейронов коры. По данным клиники, у людей с патологическим раздражением таламических структур возникают очень сильные болевые ощущения.
Интегративная функция таламуса связана с взаимодействием в нем афферентации из разных сенсорных систем. После интегративного взаимодействия афферентные влияния из таламуса направляются не только в проекционные и ассоциативные области коры, но и в базальные ганглии, гипоталамус, гиппокамп, ядра миндалевидного комплекса. Внутриталамичес-кие связи обеспечивают интеграцию сложных двигательных реакций с вегетативными процессами, регулируемыми структурами лимбической системы. Таламические интегративные механизмы имеют большое значение для условнорефлекторной деятельности, формирования эмоциональных реакций и мотива-ционного поведения. Оказывая большое влияние на состояние и деятельность коры больших полушарий, таламус сам находится под регулирующим и корригирующим влиянием коры. Эти нисходящие влияния осуществляются через прямые кортикоталамические пути и кортикоретикулоталамические связи и обеспечивают влияние коры на таламический уровень сенсорного анализа и интеграции. Множественные восходящие из таламуса в кору и нисходящие из коры в таламус пути лежат в основе деятельности единой таламокортикальной системы.
5.3.2. Гипоталамус
Гипоталамус располагается на основании мозга, образуя вентральную часть промежуточного мозга, подбугорье. Он представляет сложно организованную структуру, тесно связанную со многими отделами центральной нервной системы и гипофизом.
Гипоталамическая область является высшим подкорковым центром интеграции вегетативных, эмоциональных и моторных компонентов сложных реакций адаптивного поведения и поддержания гомеоста-
за внутренней среды организма. В гипоталамическую область включают также лежащую спереди от перекреста зрительных нервов преоптическую область и расположенное латерально субталамическое ядро Люиса.
5.3.2.1. Структура и эволюгщя
В гипоталамической области человека разные авторы выделяют от 15 до 48 ядер, которые подразделены на пять групп: преоптическую, переднюю, среднюю, наружную и заднюю. Преоптическая группа содержит медиальное и латеральное преоптические ядра и пери-вентикулярное ядро. В переднюю группу входят суп-раоптическое, супрахиазматическое и паравентрику-лярные ядра. Среднюю группу составляют вентроме-диальное, дорсомедиальное и аркуатные ядра. К наружной группе относятся латеральное гипоталамичес-кое ядро и серый бугор. В заднюю группу входят пре-мамиллярное, супрамамиллярное, латеральное и медиальное мамиллярные ядра, заднее гипоталамичес-кое ядро и субталамическое ядро Монса.
В ходе эволюции гипоталамус развивался и дифференцировался раньше, чем таламус. У круглоротых он уже является выраженной структурой, над которой надстраиваются образования архи- и палеокор-текса. У амфибий структуры гипоталамуса связаны преимущественно с примордиальной периформной корой и примордиальным гиппокампом. Более высокого уровня развития достигает гипоталамус у рептилий: в переднем отделе формируются зачатки "супра-оптического и паравентрикулярных ядер, в среднем — вентромедиального ядра. У птиц хорошо выражена дифференциация гипоталамических ядер, у млекопитающих большого развития достигает серое вещество гипоталамуса, нейроны которого' внутри ядер образу-
ют поля. Наиболее древними структурами гипоталамуса являются преоптическая и передняя группы, наиболее молодыми — серый бугор и мамиллярные тела. В онтогенезе раньше закладываются и оформляются супраоптические и паравентрикулярные ядра по сравнению с филогенетически более молодыми латеральными гипоталамическими и мамиллярными ядрами. У новорожденных гипоталамус еще полностью не сформирован, архитектоническое его формирование завершается к 3-4 годам жизни. К 13-14 годам происходит усиление гипоталамо-гипофизарных связей и нейросекреторных влияний.
Гипоталамус тесно связан афферентными и эфферентными связями со многими образованиями головного мозга. Афферентные пути связывают гипоталамус с обонятельной луковицей и обонятельным бугорком, миндалиной, хвостатым ядром, гиппокампом, таламусом, орбитальной, височной и теменной областями коры. Ведущими источниками афферентации гипоталамуса являются структуры лимбической системы переднего мозга и ретикулярной формации мозгового ствола. Гипоталамус является для гиппокампа таким же ключевым подкорковым центром, как дорсальный таламус для новой коры. Эфферентные пути из гипоталамуса идут к таламусу, ретикулярной формации ствола и вегетативным центрам среднего, продолговатого и спинного мозга (паравентрикулярная система). Особая система эфферентных путей тесно связывает гипоталамус с адено- и нейрогипофизом, образуя гипоталамо-гипофизарную систему. Постоянство связей гипоталамуса у животных разного уровня развития свидетельствует о том, что его деятельность связана с регуляцией таких функций организма, которые не претерпели существенных изменений в ходе эволюции.
5.3.2.2. Особенности нейронных, систем
Нейронные системы гипоталамуса обеспечивают управление всеми вегетативными функциями организма и поддержание постоянства его внутренней среды. В связи с выполнением этих функций они обладают рядом особенностей: 1) непосредственной химической или физико-химической чувствительностью к содержимому крови или его изменению; 2) исключительной васкуляризацией этого отдела мозга и отсутствием здесь гематоэнцефалического барьера, отделяющего клетки мозга от кровеносных сосудов; 3) способностью нейронов некоторых ядер (супраоптического, па-равентрикулярного) секретировать пептиды, нейромо-дуляторы, вещества, активирующие работу гипофиза, таким образом осуществляя участие в регуляции работы эндокринной системы; 4) возможностью эффективного контроля за деятельностью симпатической и парасимпатической вегетативных нервных систем. Это — способности нейронных систем гипоталамуса улавливать изменения внутренней среды и адекватно реагировать, влияя на вегетативные функции организма гуморальным путем через эндокринную систему и нервным путем через симпатическую и парасимпатическую системы.
Показано, что раздражение наружной и задней группы ядер приводит к повышению тонуса симпатической нервной системы. Стимуляция серого бугра и некоторых ядер средней группы, наоборот, вызывает снижение тонуса этой системы. Воздействие на ряд ядер передней группы приводит к парасимпатическим реакциям. Гесс (1954) считал, что в гипоталамусе имеются две антагонистические зоны: эрготроп-ная, или симпатическая, расположенная в ядрах задней группы и области околоводопроводного серого вещества, и трофотропная, или парасимпатическая,
локализованная в передних отделах гипоталамуса, преоптической области. Однако в настоящее время показано, что имеет место мозаичное распределение и перекрытие участков, активизирующих симпатическую и парасимпатическую системы по всему гипоталамусу с некоторым преобладанием первых в задних отделах, а вторых — в передних. Гипоталамичес-кие структуры совместно с рядом других образований ЦНС участвуют в организации целостных веге-топсихомоторных реакций, которые могут содержать одновременно как симпатические, так и парасимпатические компоненты.
5.3.2.3. Центры гипоталамуса
В гипоталамусе расположены важнейшие центры, организующие поддержание постоянства внутренней среды (гомеостаз) и приспособление внутренней среды к изменению условий жизнедеятельности (гомео-кинез).
Центр теплорегуляции включает в себя структуры, регулирующие теплообразование, и структуры, регулирующие теплоотдачу. Раздражение ядер задней группы приводит к повышению температуры в результате повышения теплопродукции за счет усиления обменных процессов и дрожания скелетной мускулатуры. Стимуляция паравентрикулярных ядер приводит к изменению теплоотдачи в результате усиления потоотделения, расширения просвета кожных сосудов, а также к торможению мышечного дрожания. Нейроны центра терморегуляции чувствительны к изменениям температуры притекающей к ним крови.
Центр голода и насыщения, регулирующий сложное пищевое поведение, связан с нейронными группировками в средних и наружных ядрах. Нейроны центра голода возбуждаются при снижении содержа-
ния питательных веществ (глюкозы, аминокислот, жирных кислот) в крови. Нейроны центра насыщения возбуждаются при достаточном повышении содержания в крови питательных веществ или каких-то особых веществ, выделяемых пищеварительным трактом при его деятельности. Чувствительность отдельных нейронных группировок этого центра к «голодной» или «сытой» крови лежит в основе запуска сложного пищевого поведения и его прекращения.
Центр жажды и ее удовлетворения, расположенный в гипоталамусе, организован сходным образом. Стимуляция структур, расположенных кнаружи от супраоптического ядра, ведет к резкому увеличению потребления воды, разрушение этих структур — к почти полному отказу от воды. Возбуждение центра жажды связано с чувствительностью его нейронов к повышению осмотического давления крови при недостатке воды в организме. Это включает целый комплекс сложных поведенческих реакций поиска воды и питья, направленных на снижение осмотического давления крови, омывающей мозг, до нормальной величины. Возбуждение центров голода и жажды сопровождается соответствующими субъективными переживаниями.
Центр регуляции полового поведения, расположенный в гипоталамусе, участвует в регуляции комплекса функций, связанных с размножением. Изолированное разрушение области серого бугра ведет к атрофии половых желез. Известно немало клинических случаев быстрого полового созревания при возникновении опухоли в области серого бугра. При повреждениях средних областей гипоталамуса описаны случаи превращения вторичных мужских половых признаков в женские. У примерно половины больных с патологией гипоталамуса наблюдались нарушения
функций половой системы. Результаты экспериментов и клиники свидетельствуют, что структуры передних отделов гипоталамуса оказывают ускоряющее действие на половое развитие, задней части — замедляющее действие. Опыты с вживлением электродов в эти структуры гипоталамуса, при которых животным предоставлена возможность самим, нажимая на педаль, включать стимулирующий ток, показали, что животные стремятся проводить самораздражение с максимальной частотой и продолжительностью (Олдс, 1954). Очевидно, что при таком раздражении у животных возникают приятные им реакции; при случайном раздражении в ходе нейрохирургических операций пациенты испытывали приятные ощущения. Локализованный в заднем гипоталамусе центр удовольствия, взаимодействуя с нейронными образованиями других структур лимбической системы, принимает участие в организации эмоциональной сферы полового поведения. При раздражении структур переднего гипоталамуса у животных возникают реакции страха и ярости, что свидетельствует о существовании центра неудовольствия, связанного с включением отрицательных эмоций. Гипоталамическая область вместе с рядом структур конечного мозга играет важную роль в организации сложнейших биологических рефлексов, направленных на обеспечение основных биологических функций, формирование соответствующих мотиваций. Удовлетворение мотиваций сопровождается положительным эмоциональным состоянием, неудовлетворение — отрицательным.
В гипоталамусе имеются структуры, принимающие участие в регуляции чередования бодрствования и сна — одного из центров регуляции цикла бодрствование — сон. Раздражение латеральной базальной пре-оптической области мозга у животных обладает вы-
раженным гипногенным действием. Стимуляция этой области вызывает развитие сна или изменение электрической активности, характерной для сна; разрушение нарушает развитие сна. У людей поражение гипоталамуса часто сопровождается нарушениями сна и изменениями ЭЭГ, характерными для сна. Переход от сна к бодрствованию и наоборот сопровождается комплексом изменений всех вегетативных процессов. Во время сна преобладает тонус парасимпатической системы, во время бодрствования — симпатической. Супрахиазматическое ядро гипоталамуса — важнейшее звено в организации биоритмов, центрального механизма «биологических* часов, организующих суточные циклы.
5.3.2.4. Регуляция работы гипофиза
Особая роль гипоталамуса в регуляции вегетативных функций организма определяется тем, что, наряду с регуляцией вегетативной нервной системы, он регулирует работу гипофиза, а через него и всей эндокринной системы. Между гипоталамусом и гипофизом существуют тесные структурно-функциональные связи, объединяющие их в гипоталамо-гипофи-зарную систему (рис. 5.4). Задняя доля гипофиза (ней-рогипофиз) связана с супраоптическим ядром гипоталамуса прямыми нервными связями: супрастино-ги-пофизный тракт представляет собой аксоны клеток супраоптического ядра. Нейросекрет, вырабатываемый нейронами гипоталамуса, по аксонам транспортируется в заднюю долю гипофиза и выделяется в кровь на окончаниях аксонов при их возбуждении, как медиатор. Основными гормонами задней доли гипофиза являются антидиуретический гормон — вазо-прессин, регулирующий водный обмен, и окситоцин, регулирующий мускулатуру матки. С передней долей
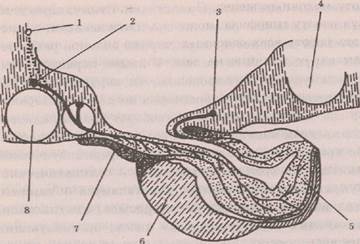
Рас. 5.4. Схема, показывающая связи гипоталамуса с передним и. задним гипофизом: 1 — паравенпгрикулярное ядро, 2 — суп-раоптическое ядро, 3 — бугрогипофизарный тракт, 4 — сосковидное тело, 5 — задняя доля гипофиза — нейрогипофиз, 6 — передняя доля гипофиза — аденогипофиз, 7 — сосуды гипофи-зарнои ножки, 8 — зрительный, перекрест (Загер, 1962)
гипофиза (аденогипофизом) гипоталамус связан через общую воротную сосудистую систему. Попадающие в кровеносные сосуды продукты нейросекреции гипоталамуса с током крови — нейрогуморальным путем -регулируют работу передней доли гипофиза, которая вырабатывает тройные гормоны, контролирующие работу других эндокринных желез. Важнейшие из них: адренокортикотропный гормон (АКТГ) стимулирует выделение гормонов коры надпочечников; тирео-тропный гормон (ТТГ) стимулирует работу щитовидной железы; фолликулстимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ) регулируют работу половых желез. Кроме них в передней доле гипофиза вырабатывается гормон роста — соматотропный гор-Мрн (CTF) и гормон пролактин, стимулирующий ра-
боту молочных желез. Поэтому гипоталамус, регулируя работу гипофиза, может эффективно контролировать работу всех основных звеньев эндокринной системы и ее влияние на вегетативные функции. Так, путь гипоталамус — гипофиз — надпочечники является важнейшим для изменения вегетативных функций и состояния организма при стрессе.
Таким образом, тесные связи гипоталамуса со многими структурами головного мозга, контроль над вегетативной нервной и эндокринной системами, многообразие реакций и клинических проявлений при его раздражении и поражении отражают роль гипоталамуса в интегративной деятельности мозга. В настоящее время гипоталамус рассматривают не как отдельную изолированную часть мозга с большим числом специализированных центров, а как важнейшее звено лимбико-ретикулярного комплекса, интегрирующего вегетативные, эмоциональные и моторные компоненты адаптивного поведения.
5.4. Физиология мозжечка.
5.4.1. Анатомо-физгюлогические особенности внутримозжечковых связей
Мозжечок в филогенезе развивается как орган, обеспечивающий рефлекторные реакции животных, связанные с преодолением влияния тяжести и инерции при перемещении в пространстве. Помимо этого, так как мозжечок имеет представительства сенсорных и вегетативных центров, его деятельность отражается также и на этих функциях.
Впервые мозжечок появляется у круглоротых и наибольшего развития достигает у птиц и млекопитающих. У амфибий происходит количественная редукция всех структур мозжечка, связанная с переходом
животного из водной среды на сушу. Особенно четко редукция выражена у бесхвостых амфибий. Считается, что причина такой редукции обусловлена сужением и выпадением афферентных связей мозжечка с органом боковой линии, играющей первостепенную роль в локомоции рыб. Морфологические и функциональные особенности развитого мозжечка у птиц связаны с их передвижением в воздушной среде, т.е. в безопорном пространстве. У птиц мозжечок включает три доли: тело, или переднюю долю, — гомолог язычка, центральной дольки и вершинки млекопитающих; флоккуло-нодулярную долю — эквивалент втулочки, узелка и пирамидки мозжечка млекопитающих; среднюю долю, филогенетически более молодую, соответствующую листку и простой дольке млекопитающих. Мозжечок млекопитающих обладает существенным морфологическим дополнением, отличающим его от всех нижестоящих классов позвоночных. Усиленный рост его боковых формаций, происходивший параллельно с быстрым развитием другой филогенетически молодой структуры — паллиума, привел к образованию четко выраженных полушарных экранных структур.
Date: 2016-11-17; view: 335; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |