Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Мал. 2.4. Механізм виникнення двофазного потенціалу дії: а, б — відвідні електроди
При внутрішньоклітинному відведені можна кількісно охарактеризувати зміни МПС при генерації ПД. У типовому внутрішньоклітинному ПД розрізняють пік (верхівку), висхідну, низхідну фази і слідові потенціали. Встановлено, що під час висхідної фази ПД відбувається не просто зникнення трансмембранної різниці потенціалів (деполяризація мембрани), а виникає різниця потенціалів протилежного знака (овершут), внутрішня частина мембрани в ділянці генерації ПД стає на певний час зарядженою позитивно відносно зовнішньої частини мембрани, зарядженої негативно.
Під час низхідної фази ПД відновлюється початкове значення МП (реполяризація мембрани). Тривалість піку ПД різних збудливих тканин варіює від 0,5 до 3 мс. Зміни поляризації мембрани, які спостерігаються після завершення високовольтної частини ПД (піку), називають слідовими потенціалами. Розрізняють два види слідових потенціалів — слідову деполяризацію (слідовий негативний потенціал) і слідову гіперполяризацію (слідовий позитивний потенціал), амплітуда яких не перевищує кількох мілівольт (5-10% рівня піку), а тривалість буває різною — від кількох мілісекунд до десятків секунд.
Іонний механізм генерації ПД. В основі генерації ПД лежать зміни проникності клітинної мембрани, що виникають послідовно за часом. У момент подразнення провідність мембрани для Nа+ зростає. Nа+ починає надходити в клітину, знижуючи внутрішній негативний заряд мембрани, тобто виникає деполяризація клітинної мембрани. При досягненні мембраною певного — критичного — рівня деполяризації виникає лавиноподібний потік Nа+ всередину клітини, що зумовлює появу висхідного коліна ПД. Заряд мембрани в місці подразнення змінюється — виникає реверсія знака МП.
Натрієва провідність досягає максимуму менш ніж за 1 мс після початку ПД, а потім різко знижується, зникаючи зовсім протягом 0,5 мс, хоча деполяризація мембрани ще зберігається. Це явище називають натрієвою інактивацією, яка існує доти, доки триває деполяризація мембрани. Натрієва активаційна система відновлюється, коли МПС повертається до початкового рівня або навіть при певній гіперполяризації мембрани
Зміни збудливості клітини під час збудження. Процес збудження проходить ряд фаз, під час яких змінюється збудливість тканини. Поява місцевого збудження, що не поширюється, характеризується невеличким, дуже коротким за часом, підвищенням збудливості. При переході місцевого збудження в таке, що поширюється, виникає пік потенціалу дії, обумовлений переходом іонів натрію всередину клітини, і збудливість тканини різко падає. Стан тканини, коли вона після подразнення тимчасово не реагує на повторне подразнення будь-якої сили, називається абсолютною рефрактерністю. Потім протягом кількох мілісекунд після закінчення абсолютної рефрактерності нове збудження можна викликати лише більшою силою подразнення, ніж перший ПД. Значення ПД в цей час також зменшене, оскільки натрієва система не повністю відновлена після інактивації під час першого ПД. Цей відрізок часу, протягом якого відбувається відновлення збудливості клітини, називають відносним рефрактерним періодом.
Абсолютний рефрактерний період обмежує максимальну частоту генерації ПД живою клітиною. Оскільки у більшості нервових клітин ПД триває близько 2 мс, то максимально можлива частота генерації ПД у них буде 500 за 1 с. Проте є клітини із ще коротшим рефрактерним періодом, які мають частоту генерації до 1000 за 1 с. Зазвичай частота генерації ПД в ЦНС не перевищує кількох десятків ПД за 1 с.
Після фази абсолютної рефрактерності збудливість тканини поступово відновляється до вихідного рівня. Цей період зниженої збудливості одержав назву відносної рефрактерності. Фаза відносної рефрактерності збігається із переходом піку в негативний слідовий потенціал. Відносна рефрактерність продовжується в нервових волокнах від 0,001 – 0,01 с, а в м'язах до 0,03 с. У цю фазу можна, використовуючи більш сильний подразник, викликати новий імпульс збудження. Потім наступає фаза підвищеної збудливості або екзальтаційна. Вона триває в нерві – 0,02, а в м'язі – 0,05 с.
Цей період підвищеної збудливості збігається зі слідовою електронегативністю.
Під час слідової деполяризації збудливість клітини підвищується (фаза екзальтації), а під час слідової гіперполяризації — знижується, оскільки значення МПС у цей час зростає, при цьому збільшується і критичний (пороговий) рівень деполяризації мембрани.
У живій клітині існують дві системи руху іонів крізь мембрану. Одна із них здійснюється за градієнтом концентрації іонів і не потребує витрати енергії — це пасивний іонний транспорт. Він відповідає за підтримання МПС і виникнення ПД, здійснюється через іонні канали. Друга система руху іонів крізь мембрану відбувається проти концентраційного градієнта і полягає у "викачуванні" Na+ із цитоплазми і "закачуванні" К+ всередину клітини за допомогою і онних насосів. Цей механізм можливий тільки із витратами енергії і називається активним іонним транспортом.
Функції іонних каналів. У клітинній мембрані існують пори, або канали. Терміном "канал" позначають той шлях, яким іони проходять крізь мембрану за електрохімічним градієнтом. У клітинній мембрані існує кілька різних іонних каналів: селективних та неселективних. Перші пропускають лише якийсь певний іон: Na+, К+ або Са2+, а другі — кілька іонів. Крім того, селективні канали поділяють на потенціалозалежні, потенціалонезалежні (хемочутливі) і канали витоку.
Потенціалозалежні канали. Провідність цих каналів контролюється рівнем МПС, тобто ці канали відкриваються (активуються) і закриваються (інактивуються) у відповідь на зміни МПС. Потенціалозалежні канали функціонують в електрозбудливих мембранах і беруть участь у генерації ПД. Електрична збудливість пов’язана із існуванням у таких каналах особливого молекулярного пристрою — воріт,відкривання і закривання яких визначається електричним полем мембрани.
Ворота можуть перебувати тільки у двох положеннях: повністю відчиненими чи повністю зачиненими, тому провідність поодинокого іонного каналу є сталою. Через відкриті канали іони рухаються за концентраційним і електрохімічним градієнтами. Ці потоки іонів зумовлюють зміну МПС, що, в свою чергу, змінює середню кількість відкритих каналів і відповідно рівень іонних струмів.
Потенціалозалежними є натрієві, калієві і кальцієві канали. Натрієві і кальцієві канали відповідають за ранній (початковий) струм вхідного (в клітину) напрямку, а калієві — за пізній (відставлений) струм вихідного (з клітини) напрямку.
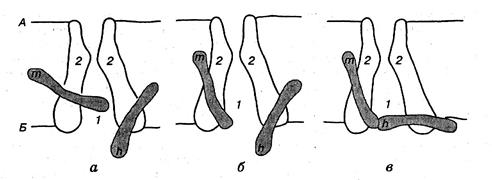 |
Натрієвим каналам властива досить висока, але не абсолютна вибірковість Nа+, оскільки більшою або меншою вони є проникними також для іонів літію, талію і навіть деяких органічних катіонів (гідразин, амоній). Отвір натрієвого каналу має прямокутну форму розмірами 0,3х0,3 нм. Вузька частина каналу, що є селективним фільтром, дуже коротка і має одиничний негативний заряд. Цей заряд притягує катіони, відштовхує аніони, забезпечуючи вибіркову проникність для Nа+ і подібних до нього катіонів (мал 2.5).
Мал. 2.5.Потенціалозалежний натрієвий канал:
Внутрішня частина каналу (1), утвореного макромолекулою білка (2), — це селективний фільтр; т — активаційні. h — інактиваційні: - ворота каналу; а — стан спокою (активаційні ворота зачинені, інактнваційні — відчинені); б — початок деполяризації мембрани (швидке відчинення активаційних воріт); в — інактивація каналу (інактиваційні ворота зачинені); поверхня клітинної мембрани: А — зовнішня; Б — цитоплазматична
Активаційні та інактиваційні ворота розміщені біля внутрішнього кінця натрієвого каналу, причому інактиваційні ворота спрямовані в бік цитоплазми. У стані спокою активаційні ворота закриті, а інактиваційні відкриті. Під час деполяризації мембрани спочатку відкриті обидва типи воріт, тобто канал пропускає Nа+. Потім інактиваційні ворота повільно закриваються — канал інактивується. Лише після закінчення деполяризації поступово відкриваються іиактиваційні ворота, а активаційні закриваються і канал повертається до початкового стану.
Деякі речовини специфічно змінюють процеси активації та інактивації натрієвих каналів. Так, батрахотоксин спричинює стійку деполяризацію клітинної мембрани, підвищуючи gNa.Він повністю усуває процес натрієвої інактивації так, що натрієві канали стають постійно відкритими. Тетродотаксин дуже швидко і різко пригнічує ранній іонний струм крізь мембрану, але тільки коли він діє із зовні, тобто на чутливі до тетродотоксину рецепторні структури натрієвих каналів, розміщені на зовнішній поверхні мембрани.
Калієві канали за будовою подібні до натрієвих, але відрізняються своєю вибірковістю, спрямованістю потоку іонів, кінетикою процесів інактивації і активації. В усіх збудливих клітинах калієві канали відповідають за пізній іонний струм. Діаметр калієвого каналу у найвужчій його частині становить 0,3 нм. Отже, калієві канали проникні для катіонів розміром 0,26-0,З нм (К+,RЬ+).
Активація цих каналів порівняно із натрієвими відбувається досить повільно. Крім того, протягом перших 10 мс деполяризації немає ніякої калієвої інактивації, вона виникає тільки під час дуже тривалої (кілька секунд) деполяризації мембрани.
Викладене вище про співвідношення між процесами активації та інактивації калієвих каналів властиве лише нервовим волокнам. У мембрані багатьох нервових і м'язових клітин існують калієві канали, які інактивуються порівняно швидко. Виявлено також калієві канали із швидкою активацією. Нарешті, є калієві канали, які активуються не мембранним потенціалом, а внутрішньоклітинним Са2+. Щільність розміщення калієвих каналів на мембрані дещо менша, ніж натрієвих. Специфічним блокатором калієвих каналів є тетраетиламоній (ТЕА), і они гідрогену, амінопіридини. ТЕА діє із будь-якого боку клітинної мембрани.
Кальцієві канали. Ретельне дослідження функції йонних каналів різних нервових клітин дало змогу модифікувати йонну модель генерації ПД, запропоновану Ходжкіним і Хакслі, до якої входили лише натрієві й калієві канали. Виявилося, що Са2+ також беруть активну участь у генерації ПД. З'ясувалося також, що в деяких клітинах вхідний струм створюється переважно Са2+ (нейрони молюсків, гладком'язові клітини). Крім порівняно швидкого вхідного кальцієвого струму, що бере участь у генерації ПД, було виявлено ще один дуже повільний кальцієвий струм. Цей струм відповідає за повільну деполяризацію (протягом кількох секунд), яка зумовлює генерацію серій імпульсів деяких пейсмекерних клітин, наприклад серцевих провідних міоцитів (клітин Пуркіньє).
Селективність (вибірковість) кальцієвих каналів зумовлена наявністю в ділянці зовнішнього входу хімічних груп, які мають підвищену спорідненість до двовалентних катіонів: іони кальцію зв'язуються із такими групами і тільки після цього проходять до порожнини каналу. До деяких двовалентних катіонів, наприклад Мn2+, спорідненість цих хімічних груп така велика, що, зв'язуючись із ними, ці катіони блокують рух Са2+ через канал. Специфічними блокаторами кальцієвих каналів є харибдотоксин, вилучений із отрути скорпіона, а також такі лікарські речовини, як верапаміл та ніфедипін.
Характерною особливістю кальцієвих каналів є залежність їх від клітинного метаболізму, зокрема від циклічних нуклеотидів (цАМФ, цГМФ), які регулюють процеси фосфорилування і дефосфорилування білків кальцієвих каналів. Йони кальцію ефективно впливають на синтез білків і забезпечують транспорт утворених молекул до аксонів і дендритів. Відкриття цього явища безпосередньо свідчило про те, що процеси на мембрані прямо пов'язані із процесами всередині клітини.
Дослідження останніх років показали, що Са2+ є важливим регулятором активності збудливих систем. Переважна кількість Са2+, що потрапляє в клітини під час збудження, зв'язується буферними системами цитозолю, мітохондріями та іншими внутрішньоклітинними депо. Проте та невелика частина Са2*, що залишається вільною, бере активну участь у таких процесах, як регуляція функції К+ і Сl— каналів, Са2+, Nа+ обміну тощо.
Виявлено ще одну особливість кальцієвих каналів: вони здатні дуже специфічно реагувати на деякі фізіологічні речовини, що утворюються іншими клітинами і виділяються у внутрішнє середовище організму (катехоламіни, пептиди тощо). Під впливом цих речовин канали активуються чи, навпаки, інактивуються
Внаслідок цього змінюється активність клітини.
Потенціалонезалежні (хемочутливі) канали. Активність цих каналів контролюється тільки хеморецепторами. Активація хеморецепторів медіаторами супроводжується здебільшого відкриванням каналів. Хемочутливі потенціалонезалежні канали функціонують у субсінаптичних мембранах і беруть участь у: генерації синоптичних потенціалів. Вони реагують не тільки на нейромедіатори, а й на гормонита деякі інші фізіологічно активні речовини.
Канали витоку також потенціалонезалежні, але вони не контролюються хеморецепторами. Ці канали відіграють істотну роль у сумарній провідності мембран у стані спокою, саме їхньою активністю пояснюють менші значення МПС порівняно із розрахованим за рівнянням. Функцію цих каналів ще не з’ясовано.
Кожний потенціалозалежний іонний канал навіть за сталих значень МПС то відкривається, то закривається. Це відбувається незакономірно, тобто процес має ймовірний (стохастичний) характер. Таке явище викликає переміщення іонів і створює електричний шум.
У цей ж час із клітини виходить незначна кількість іонів калію. За один імпульс клітина втрачає мільйонну частку всієї кількості калію. У результаті іонних переміщень настає деполяризація – клітинний потенціал падає до нуля. Потім відбувається перезарядження мембрани – її реполяризація. Через якийсь час потенціал мембрани відновляється і клітина знову може реагувати на подразнення.
Переміщення іонів натрію всередину клітини є причиною появи місцевого колового електричного струму, який поширюється по поверхні мембрани. Всередині клітини круговий струм йде від збудженої ділянки до незбудженої, а зовні – від місця спокою до збудженої ділянки. Проникаючи в сусідні незбуджені ділянки, місцевий струм відчиняє пори для натрію, створюючи тут новий імпульс. Зароджуючись у кожній наступній точці нервового або м'язового волокна, імпульс поширюється на значну відстань.
Історія вчення про біоструми нараховує вже декілька сторіч. Вперше людина спостерігала біоструми у електричних риб. У 1791 році італійський фізіолог Луїджі Гальвані довів, що в живій тканині є біоструми. Наявність біострумів він довів дослідами, які ввійшли в історію за назвою 1 і 2 дослідів Гальвані. Він установив наявність так званих струмів спокою. Під час збудження на поверхні мембрани реєструються струми дії. Між ними є суттєва різниця. Якщо струми спокою однофазні і реєструються довгостроково, то струми дії двофазні і короткочасні.
У зв'язку зі зміною потенціалу дії, а отже і із тривалістю проходження фаз збудливості, знаходиться і функціональна рухливість або лабільність тканини.
Вчення про лабільність створив М. Є. Введенський. Для визначення частоти виникаючих потенціалів дії, він у 1884 році застосував слухавку, сполучену із нервом нервово – м'язового препарату. Під час подразнення нерва із різною частотою в слухавці виникали звуки – тони різної висоти. Чим частіше задавалося подразнення, тим вище був тон. За граничною висотою тону можна було судити про максимальну кількість відтворених за секунду імпульсів збудження. Таке ж положення спостерігалося і під час подразнення м'язів.
На підставі дослідів М. Є. Введенський створив вчення про функціональну рухливість, або лабільність, що розкриває закономірності ритмічного збудження, що поширюється.
Основним у вченні про лабільність є положення про те, що в кожній тканині поодинокий імпульс збудження продовжується визначений час. Тому М. Є. Введенський визначав лабільність, як швидкість, із якою у тканині виникає і встигає закінчитися повний період окремого імпульсу збудження. Тривалість окремого імпульсу збудження, що вимірюється за тривалістю фази абсолютної рефрактерності, характеризує тканину або орган із боку швидкості біохімічних і фізіологічних процесів, які відбуваються в них. Чим швидше відбувається перебіг процесів, що складають окремий імпульс збудження, тим вище лабільність і, отже, більше окремих імпульсів збудження може виникнути в тканині за визначений проміжок часу. Для виміру лабільності був запропонований показник – міра лабільності.
Міра лабільності – те максимальне число імпульсів збудження, що виникає за 1с у відповідь на таке ж максимальне число подразнень.
Високу лабільність мають мієлінові соматичні нерви, менш високу – безмієлінові вегетативні нерви. Максимальний ритм збудження для мієлінового нерва – 500, для вегетативного – 200 імпульсів за секунду. Лабільність скелетних м'язів вище, ніж непосмугованих. Так, максимальний ритм збудження для скелетних м'язів становив у середньому 200 імпульсів за секунду, а для непосмугованих – у десятки разів менше.
Лабільність у процесі росту і розвитку організму збільшується, а під час старіння – зменшується. Вона не залишається постійною і за різних функціональних змін в організмі, тому що стомлення або розвиток патологічних процесів знижує лабільність.
Під час ритмічного подразнення із визначеною частою тканина не відразу відповідає відповідною кількістю імпульсів збудження. Це залежить від того, що лабільність до початку подразнення відрізняється від лабільності під час подразнення – вона змінюється в зв'язку із діяльністю. Дія подразника і виникаючі імпульси збудження змінюють швидкість процесів обміну речовин у тканині. Остання набуває спроможності відтворювати більш частий, раніше недоступний їй ритм.
Дослідами М. Є. Введенського вперше було встановлено, що найкраще скорочення м'яза спостерігається при нанесенні на нерв нервово- м'язового препарату подразнень у більш рідкому ритмі. Такий найбільше вигідний у робочому відношенні ритм подразнення був названий оптимальним. За оптимального ритму подразнень кожний новий імпульс збудження виникає під час фази екзальтації, створеної попереднім імпульсом, у результаті чого і відбувається максимальне скорочення м'яза. Оптимальний ритм збуджень менше максимального у декілька разів. Наприклад, у жаби максимальний ритм для рухового нерва складає 500 імпульсів за секунду, а оптимальний – 100 – 150, для литкового м'яза відповідно – 120 – 200 і 30 – 50 імпульсів за секунду. При дуже частих подразненнях, що перевищують оптимальний ритм, скорочення м'яза зменшується і навіть зовсім припиняється – спостерігається песимум ритму подразнення або песимум частоти. Він спостерігається в тому випадку, коли частота подразнень перевищує міру лабільності. Під впливом першого подразнення тканина збуджується, імпульс збудження ще не закінчився, а вже наноситься повторне подразнення, яке діє під час фази абсолютної рефрактерності. Хоча те подразнення не в змозі викликати збудження, воно усе ж не залишається без наслідків – поглиблює фазу абсолютної рефрактерності і створює утруднення для дії наступних за ним подразнень.
Тому кожне нове подразнення, що посилається в дуже частому ритмі, знижує лабільність тканини і викликає не збудження, а якісно новий стан – гальмування.
Явища оптимуму і песимуму, відкриті М. Є. Введенським при вивченні реакцій нервово-м'язового препарату, виявилися загальнобіологічними властивостями, що спостерігаються у живій тканині у відповідь на будь-які фізичні або хімічні подразники. Встановлено, що помірні подразнення викликають оптимальні (збудливі) дії, а подразнення великої сили, частоти і тривалості песимально (гальмуюче) діють на реакції різних органів як за безпосереднього, так і за рефлекторного впливу на них.
Явища оптимуму і песимуму відзначаються не тільки в рухових реакціях, але й у реакціях секреторних органів і серцево-судинної системи. Ці явища були виявлені й в умовно-рефлекторних реакціях різної складності. Вивчаючи вплив різноманітних тривалих хімічних і фізичних подразнень на нерв нервово-м'язового препарату, М. Є. Введенський встановив закономірні стадії зміни функціонального стану нерва в ділянці, на яку наноситься подразнення.
Висока лабільність тканини забезпечує проведення імпульсів збудження без зміни, зниження лабільності трансформує часті імпульси в більш рідкісні, що можна вважати за часткове гальмування.
Чим більше знижена лабільність, чим частіше і сильніше подразнення, тим швидше настає такий стан, коли збудження, яке поширюється переходить у місцеве збудження, що не поширюється і викликає гальмування.
М. Є. Введенський вважав гальмування модифікацією збудження, яке не поширюється, стає місцевим, стійким. Гальмування являють собою наростаючі кількісні зміни за розвитку процесу збудження, що швидко переходять у якісно новий стан – стан гальмування.
Збудження характерне для таких тканин як нервова, м’язова та залозиста. Для кожної тканини розвиток збудження має свої особливості. Крім того в організмі риб є специфічні тільки для цього класу тварин електричні органи, які мають клітини, що можуть генерувати електричний струм. Тканини, із яких вони створюються, різні за походженням. Вони розвиваються із нервової, м’язової або залозистої тканини, але функція їх в організмі подібна.
Запитання для самоперевірки
1. Що таке подразливість?
2. Як класифікуються подразники?
3. Що таке збудливість?
4. Які тканини є збудливими?
5. Що таке потенціал спокою?
6. Що таке потенціал дії, як він виникає?
7. На основі яких показників вимірюється збудливість?
8. Від яких факторів залежить збудливість тканин?
9. Що таке гранична сила подразника (поріг сили)?
10. Що таке хронаксія?
11. Що таке рефрактерний період?
12. Яка буває рефрактерність?
13. Що таке лабільність?
14. Як в тканині виникає гальмування?
Date: 2016-11-17; view: 430; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |