Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Строение и функции лизосом
Лизосомы— это пузырьки, Имеющие одномембранную оболочку, которая снаружи иногда бывает покрыта волокнистым белковым слоем. Содержат набор ферментов (кислых гидролаз), которые осуществляют при низких значениях рН гидролитическое (в присутствии воды) расщепление веществ (нуклеиновых кислот, белков, жиров, углеводов). Основная функция — внутриклеточное переваривание различных химических соединений и клеточных структур.
Выделяют первичные (неактивные) и вторичные лизосомы (в них протекает процесс переваривания). Вторичные лизосомы образуются из первичных. Они подразделяются на гетеролизосомы и аутолизосомы.
В гетеролизосомах (или фаголизосомах) протекает процесс переваривания материала, который поступает в клетку извне путем активного транспорта (пиноцитоза и фагоцитоза).
В аутолизосомах (или цитолизосомах) подвергаются разрушению собственные клеточные структуры, которые завершили свою жизнь.
Вторичные лизосомы, которые уже перестали переваривать материал, называются остаточными тельцами. В них нет гидролаз, содержится непереваренный материал.
При нарушении целостности мембраны лизосом или при заболевании клетки гидролазы поступают внутрь клетки из лизосом и осуществляют ее самопереваривание (автолиз)
(3) ГЕТЕРОХРОМАТИН-участки хроматина, находящиеся в плотно упакованном состоянии в течение всего клеточного цикла. Интенсивно окрашиваются ядерными красителями и хорошо видны в световой микроскоп даже во время интерфазы. Гетерохроматич. р-ны хромосом реплицируются позже эухроматиновых и не транскрибируются. Различают факультативный и конститутивный (структурный) Г. Факультативный Г. присутствует только в одной из гомологичных хромосом. Пример Г. такого типа — вторая Х-хромосома у жен. особей млекопитающих, к-рая в ходе раннего эмбриогенеза инактивируется вследствие её необратимой конденсации. Структурный Г. содержится в обеих гомологичных хромосомах, локализован преим. в экспонированных участках хромосомы — в центромере, теломере, ядрышковом организаторе (во время интерфазы он располагается неподалёку от ядерной оболочки), обеднён генами, обогащен сателлитной ДНК и может инактивировать расположенные по соседству гены (т. н. эффект положения). Этот тип Г. очень вариабелен как в пределах одного вида, так и в пределах близких видов. Он может влиять на синапсис хромосом, частоту индуцированных разрывов и рекомбинацию.
Эухроматин, активный хроматин — участки хроматина, сохраняющие деспирализованное состояние элементарных дезоксирибонуклеопротеидных нитей (ДНП) в покоящемся ядре, т. е. в интерфазе (в отличие от других участков, сохраняющих спирализованное состояние — гетерохроматина
).Эухроматин отличается от гетерохроматина также способностью к интенсивному синтезу рибонуклеиновой кислоты (РНК) и б о льшим содержанием негистоновых белков. В нём, помимо ДНП, имеются рибонуклеопротеидные частицы (РНП-гранулы) диаметром 200—500, которые служат для завершения созревания РНК и переноса ее в цитоплазму. Эухроматин содержит большинство структурных генов организма.(4)-РНК. РНК не имеет жесткой специфической структуры и ее полинуклеотидная цепь образует изогнутые петли. В нерабочем состоянии м-РНК собрана в складки, свернута в клубок, связана с белком; а во время функционирования цепь расправляется. Матричные РНК синтезируются на ДНК в ядре. Процесс называется транскрипция (списывание).Роль м-РНК – она несет информацию об аминокислотной последовательности (т.е. о первичной структуре) синтезируемого белка. Место каждой аминокислоты в молекуле белка закодировано определенной последовательностью нуклеотидов в цепи м-РНК, т.е. в м-РНК имеются «кодовые слова» для каждой аминокислоты – триплеты, или кодоны, или генетические коды. р-РНК Она входит в состав рибосом. Полинуклеотидная цепь р-РНК легко изгибается и укладывается вместе с белком в компактные тельца. Рибосома состоит из 2-х субъдиниц – большой и малой. В рибосоме различают 2 участка – А (аминокислотный, или участок узнавания) и Р – пептидный, здесь присоединяется п/п цепь. Эти центры расположены на контактирующих поверхностях обеих субъдиниц. Рибосомы могут свободно перемещаться в клетке, что дает возможность синтезировать белки в клетке там, где это необходимо. Рибосомы мало специфичны и могут считывать информацию с чужеродных м-РНК, вместе с м-РНК рибосомы образуют матрицу. Роль р-РНК – обуславливает количество синтезируемого белка. т-РНК Содержится в цитоплазме. Основная роль – транспорт и установка аминокислот на комплиментарном кодоне м-РНК. т-РНК специфичны к аминокислотам. В неактивном состоянии она свернута в клубочек, а в активном имеет вид трилистника (клеверного листа). В молекуле т-РНК различают несколько участков: а) акцепторный стебель с последовательностью нуклеотидов АЦЦ, к нему присоединяется аминокислота. Б) участок для присоединения к рибосоме; в) антикодон – участок, комплиментарный кодону м-РНК, который кодирует аминокислоту, присоединенную к данной т-РНК.
5. Овогенез -Процесс развития женских половых клеток, в ходе которого незрелые половые клетки – овогонии, пройдя через ряд стадий (размножения, роста, деления), превращаются в зрелые, способные к оплодотворению яйцеклетки.
Если у мужчин образование сперматозоидов с наступлением половой зрелости – процесс непрерывный, то у женщин процесс созревания яйцеклетки носит циклический характер, повторяющийся через 21 – 35 дней и носящий название менструального цикла. Характеризуется он изменениями как в строении, так и функции яичников и матки.
В овогенезе в результате первого деления мейоза образуется один овоцит 1-го порядка и одна дочерняя клетка, именуемая редукционным тельцем, для которой характерно практически полное отсутствие цитоплазмы. Это тельце, как правило, остается расположенным рядом с овоцитом.
Далее следует второе деление созревания — эквационное, протекающее как обычный митоз. Однако, в отличие от чередования митозов соматических клеток здесь отсутствует отчетливая интерфаза и клетки переходят от первого деления мейоза ко второму делению мейоза без деконденсации хроматина и удвоения содержания ДНК. В образующиеся дочерние клетки (в сперматиды при сперматогенезе и в овоциты 2-го порядка при овогенезе) расходятся хроматиды от каждой из метафазных хромосом, таким образом, клетки получают истинно гаплоидный набор генетического материала (ядерная формула сперматид и овоцитов 2-го порядка — lcln).
В результате мейоза в овогенезе из одной исходной овогонии образуется 4 дифференцированные половые клетки — овоциты. В овогенезе овоциты продолжают мейоз лишь при действии мейоз-стимулирующей субстанции. Первое деление мейоза и второе деление до стадии метафазы женская половая клетка проходит, находясь в яичнике. На стадии метафазы второго деления мейоза овоцит 2-го порядка покидает яичник (овулирует) и претерпевает ана- и телофазу в маточной трубе (превращаясь в результате активирующего влияния сперматозоидов в зрелую гаплоидную половую клетку, или яйцеклетку). Если контакта со спермиями не происходит, овоцит 2-го порядка так и не завершает мейоз и погибает. В овогенезе каждая материнская клетка при каждом делении мейоза дает лишь одну полноценную половую клетку; вторая клетка -редукционное тельце. Редукционное тельце, которое образуется в результате первого деления мейоза, при втором делении также делится (но необязательно), давая два дочерних тельца. Таким образом, в результате мейоза в овогенезе образуется лишь одна дифференцированная яйцеклетка и три редукционных тельца.
Билет№11 1)При половом размножении происходит слияние гаплоидных половых клеток. гаметы образуются в результате мейоза. У одноклеточных половой размножение в виде конъюгации и копуляции. Конъюгация- оплодотворение происходит путем обмена мигрирующими ядрами, перемещающимися из одной клетки в другую по цитоплазматическому мостику, которую образуют две особи. увеличения количества особей не происходит, но происходит обмен генетическим материалом, что обеспечивает перекомбинацию наследственных свойств. Примером конъюгации могут служить: ресничные простейшие- инфузории и некоторые водоросли. Копуляция (гаметогамия)- две различающиеся по полу клетки – гаметы- сливаются и образуют диплоидную зиготу. Формы копуляции: изогамия, анизогамия, оогамия. На Iэтапе морфологические различия между клетками отсутствуют, но физиологически различаются на муж и жен-изогамия. Изогамии встречается у жгутиконосеца политома. Анизогамия(гетерогамия)-дальнейшее усложнение связанное с дифференцировкой гамет на крупные и мелкие. Форма анизогамии, когда гаметы резко различаются наз-ся оогамией. Жен гамета –крупная, неподвижная яйцеклетка. Муж- спермотозоиды - мелкие, подвижные. Оогамия характерна для животных, высших растении. У многоклеточных партеногенез-развитие гамет без оплодотворения. (у дафнии, пчел,ящериц. По способу размножения различают естественный -норм способ характерный для нектрых организмов в природе. Искусственный- вызывается по действием раздражителей на неоплодотворенную яйцеклетку.
2) Пассивный транспорт — перенос веществ по градиенту концентрации из области высокой концентрации в область низкой, без затрат энергии (например, диффузия, осмос) Простая диффузия. Характерна для небольших нейтральных молекул (H2O, CO2, O2), а также гидрофобных низкомолекулярных органических веществ. Эти молекулы могут проходить без какого-либо взаимодействия с мембранными белками через поры или каналы мембраны до тех пор, пока будет сохраняться градиент концентрации. Облегченная диффузия. Характерна для гидрофильных молекул, которые переносятся через мембрану также по градиенту концентрации, но с помощью специальных мембранных белков - переносчиков. Все транспортные белки образуют непрерывный белковый проход через мембрану. Облегченная диффузия не требует специальных энергетических затрат за счет гидролиза АТФ. Осмос - диф-зия воды ч/з полупроницаемые мембраны.
3)двумембранные: митохондрии-палочковидная /овальная форма. Отсутствуют только у прокариот и бактерии +искл зрелые эритроциты) открыл Альтман в конце 20в. Митоходр расположены в участках, где расходуется энергия. Строение, размеры и форма очень вариабельны.строение: 2 мембраны: наружная и внутренняя. Между ними располагается межмембранное пространство. Кристы-впячивания внутри мембраны.матрикс-внутреннее пространство. В состав мембран митохондрии входит белки, липиды, ДНК и РНК (способны к самопрепродукт) ф-ции: энергетические станции клетов; участие в синтезе стероидных гормонов и отдельных липидов. биолог. Процессы в результате ктрых образуется энергия:1.окислительн. реакции две фазы:анаэробная(гликолиз)с обр-ем ПВК молочн к-ты ктрых транспортир. в митохондриях выделяется энергия для образ-я 36молекул АТФ.;аэробное 2.дефосфорелирование-расщепл АТФ 3. фосфорилирование, сопряжен процессом окисления.
Пластиды — органоиды эукариотических растений и некоторых фотосинтезирующих простейших(например у эвглены зеленой). Покрыты двойной мембраной и имеют в своём составе множество копий кольцевой ДНК. Совокупность пластид клетки образует пластидом. По окраске и выполняемой функции выделяют три основных типа пластид:
Лейкопласты — неокрашенные пластиды, как правило выполняют запасающую функцию. В лейкопластах клубней картофеля накапливается крахмал. Лейкопласты высших растений могут превращаться в хлоропласты или хромопласты.
Хромопласты — пластиды, окрашенные в жёлтый, красный, зеленый или оранжевый цвет. Окраска хромопластов связана с накоплением в них каротиноидов. Хромопласты определяют окраску осенних листьев, лепестков цветов, корнеплодов, созревших плодов.
Хлоропласты — пластиды, несущие фотосинтезирующие пигменты — хлорофиллы. Имеют зелёную окраску у высших растений, харовых и зелёных водорослей. Набор пигментов, участвующих в фотосинтезе (и, соответственно, определяющих окраску хлоропласта) различен у представителей разных таксономических отделов. Хлоропласты имеют сложную внутреннюю структуру.
4) единица транскрипции у эукариот наз-ся – транскриптон. Гены эукариот отделены друг от друга районами нетранскрибируемой ДНК (межгенными спрейсерами). У прокариот гены также отделены друг от друга спрейсерами, но они значительно короче. Строение генов экзон(информативные участки)-интронной(неинформативные) структуры.РНК-полимераза разрывает водородные связи между двумя цепочками ДНК транскриптона по правилу комплементарности на нем сначала синтезируется большая молекула проинформационной РНК, списывающая инфор как с с информативной, так и с неинформативной хоны. Процессинг-разрушение неинфор-ой части РНК. Молекула иРНК формируется посредством сплайсинга (сплавления) отдельных фрагментов ферментами лигазами. Дальше иРНК выходит из ядра, идет в рибосомы, где происходит синтез белка фермента.
5)Мито́з - наиболее распространенный способ репродукции эукариотических клеток. Биологическое значение митоза состоит в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений.
Мейоз — разновидность митоза, в результате которого из диплоидных (2п) соматических клеток половых желез образуются гаплоидные гаметы (1n). В анафазеI каждая пара хромосом ведет себя независимо от другой пары, в результате большое число новых комбинации. Кроссинговер-создает мощный дополнительный резерв наследственной изменчивости. При оплодотворении ядра гаметы сливаются, и восстанавливается диплоидный набор хромосом. Таким образом, мейоз обеспечивает сохранение постоянного для каждого вида набора хромосом и количества ДНК.
Билет.
1. Осн.положения клет.теории:
1)все жив.организмы состоят из клеток. Клетка-единица строения, размножения, функционирования. Вне клетки нет жизни.
2)Клетки всех орг-мов сходны между собой по строению и хим.составу.
3)Клетки могут размножаться только путем деления.
4)Клеточное строение всех организмов-свидетельство о едином происхождении.
2. Аппарат Гольджи — мембранная структура эукариотической клетки, органелла, в основном предназначенная для выведения веществ, синтезированных в эндоплазматическом ретикулуме, секреции веществ, образования комплексных соединений, формировании лизосом. Комплекс Гольджи представляет собой стопку дискообразных мембранных мешочков (цистерн), несколько расширенных ближе к краям, и связанную с ними систему пузырьков Гольджи. В растительных клетках обнаруживается ряд отдельных стопок (диктиосомы), в животных клетках часто содержится одна большая или несколько соединённых трубками стопок.
3. Репродукция соматических клеток: митоз, амитоз(прямое деление центромеры без формирования веретена деления-у человека в печени),эндомитоз(без деления клетки в анафазе, обр-ся полиплоидные клетки),эндоредупликация(удвоение ДНК без деления центромер, обр-ся политенные хромосомы). Биологич.значение митоза заключается в точном идентичном распределении сестринских хроматид между дочерними клетками, в результате чего поддерживается постоянство кариотипа в поколениях клеток, бесполое размножение как у одноклеточных, так и у многоклеточных.
4. Генети́ческий код — свойственный всем живым организмам способ кодирования аминокислотной последовательности белков при помощи последовательности нуклеотидов.
Свойства:-триплетность:каждый кодон включает 3 нуклеотида.
-универсальность:у всех жив.организмов генетич.код одинаковый
-специфичность:каждый триплет соответствует только одной аминокислоте
-непрерывность и неперекрываемость:считывается без пропусков
-вырожденность:некоторые аминокислоты кодируются несколькими триплетами.
5. Профаза 1 мейоза состоит из ряда стадий:лептотена, зиготена, пахитена, диплотена, диакинез.
-лептотена(спирализация хромосом-в каждой по 2 хроматиды)
-Зиготена (стадия сливающихся нитей). Происходит конъюгация гомологичных хромосом. При конъюгации образуются биваленты.
-Пахитена (стадия толстых нитей).Завершается репликация ДНК (образуется особая пахитенная ДНК). Завершается кроссинговер – перекрест хромосом, в результате которого они обмениваются участками хроматид.
-Диплотена (стадия двойных нитей). Гомологичные хромосомы в бивалентах отталкиваются друг от друга.
-Диакинез (стадия расхождения бивалентов). Хромосомы в бивалентах соединены лишь концами хроматид.
До мейоза1-2n4c,после- n2c.
Билет.
1.Основное отличие прокариотических клеток от эукариотических заключается в том, что их ДНК не организована в хромосомы и не окружена ядерной оболочкой. Эукариотические клетки устроены значительно сложнее. Их ДНК, связанная с белком, организована в хромосомы, которые располагаются в особом образовании, по сути самом крупном органоиде клетки - ядре. Кроме того, внеядерное активное содержимое такой клетки разделено на отдельные отсеки с помощью эндоплазматической сети, образованной элементарной мембраной. Эукариотические клетки обычно крупнее прокариотических. Их размеры варьируют от 10 до 100 мкм, тогда как размеры клеток прокариот (различных бактерий, цианобактерий - сине- зеленых водорослей и некоторых других организмов), как правило, не превышают 10 мкм, часто составляя 2-3 мкм. В эукариотической клетке носители генов - хромосомы - находятся в морфологически оформленном ядре, отграниченном от остальной клетки мембраной. Эукариотическая клетка имеет разнообразные постоянные внутриклеточные структуры - органоиды (органеллы), отсутствующие в прокариотической клетке.
Прокариотические клетки могут делиться на равные части перетяжкой или почковаться, т.е. образовывать дочернюю клетку меньшего размера, чем материнская, но никогда не делятся путем митоза. Клетки эукариотических организмов, напротив, делятся путем митоза (исключая некоторые очень архаичные группы). Хромосомы при этом "расщепляются" продольно (точнее, каждая нить ДНК воспроизводит около себя свое подобие), и их "половинки" - хроматиды (полноценные копии нити ДНК) расходятся группами к противоположным полюсам клетки. Каждая из образующихся затем клеток получает одинаковый набор хромосом.
Рибосомы прокариотической клетки резко отличаются от рибосом эукариот по величине. Ряд процессов, свойственных цитоплазме многих эукариотических клеток, - фагоцитоз, пиноцитоз и циклоз (вращательное движение цитоплазмы) - у прокариот не обнаружен. Прокариотической клетке в процессе обмена веществ не требуется аскорбиновая кислота, но эукариотические не могут без нее обходиться.
Прокариоты имеют двигательные приспособления в виде жгутиков или ресничек, состоящих из белка флагеллина. Двигательные приспособления подвижных эукариотических клеток получили название ундулиподиев, закрепляющихся в клетке с помощью особых телец кинетосом. Электронная микроскопия выявила структурное сходство всех ундулиподиев эукариотических организмов и резкие их отличия от жгутиков прокариот.
2. Гиалоплазма - (от греч. hyalos — стекло и плазма), основная плазма, матрикс цитоплазмы, сложная бесцветная коллоидная система в клетке, способная к обратимым переходам из золя в гель. В состав Г. входят растворимые белки (ферменты гликолиза, активации аминокислот при биосинтезе белка, многие АТФ-азы и др.), растворимые РНК, полисахариды, липиды. Через Г. идёт транспорт аминокислот, жирных к-т, нуклеотидов, Сахаров, неорганич. ионов, перенос АТФ. Состав Г. определяет буферные и осмотич. свойства клетки. Гиалоплазмой наз. таюке сильно преломляющую лучи света эктоплазму саркодовых.
Кариоплазма ядерный сок (кариоплазма, кариолимфа, нуклео-плазма), содержимое клеточного ядра, заполняющее пространство между хроматином, ядрышком и другими структурами. Содержит различные ферменты, нуклеотиды, аминокислоты и другие вещества, необходимые для обеспечения синтеза нуклеиновых кислот и субъединиц рибосом, транспортируемых из ядра в цитоплазму. В ядерном соке находятся также нитевидные белковые молекулы, составляющие ядерный матрикс, который, подобно цитоскелету в цитоплазме, выполняет в ядре роль каркаса.
3.. Клеточный цикл -это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти. Клеточный цикл составляют два периода:
1) период клеточного роста-интерфаза. идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
Интерфаза состоит из нескольких периодов: G1-фазы- фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов; S-фазы (синтетическая), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они есть). G2-фазы, во время которой идет подготовка к митозу.У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G1 фаза. Такие клетки находятся в фазе покоя G0.
2) период клеточного деления-митоз.
Период клеточного деления (фаза М) включает две стадии:
кариокинез (деление клеточного ядра);
цитокинез (деление цитоплазмы).
4. Транскрипция. Чтобы синтезировать белки с заданными функциями и свойствами к месту их построения поступает инструкция о порядке включения аминокислот в пептидную цепь. Эта инструкция заключена в нуклеотидной последовательности матричных, или информационных РНК, синтезируемых на соотвествующей участках ДНК. Процесс синтеза иРНК наз ся транскрипцией.
Синтез и РНК, начинается с обнаружения РНК-полимеразой особого участка в молекуле дНК, который указывает место начала транскрипции - промотора. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Две цепи ДНК в этом месте расходятся и на одной из них фермент осуществляет синтез мРНК. Сборка рибонуклеотидов в цепь происходит с соблюдением их комплиментарности нуклеотидам ДНК, а также антипараллельно по отношению к матричной цепи ДНК. В связи с тем, что РНК полимераза способна собирать полинуклеотид лишь от 5’ конца к 3’ концу, матрицей для транскрипции может служить только одна из двух цепей ДНК, а именно та которая обращена к ферменту своим 3’ концом. тТакую цепь наазывают кодогенной. Антипараллельность соединения двух полинуклеотидных цепей в молекуле ДНК позволяет РНК- полимеразе правильно выбрать матрицу для синтеза мРНК.
Продвигаясь вдоль кодогенной цепи ДНК, РНК-полимераза осуществляет постепенное точное переписывание информации до тех пор, пока не встречает специфическую нуклеотидную последовательность – терминатор транскрипции. В этом участке РНК-полимераза отделяется как от матрицы ДНК, так и от вновь синтезированной и РНК. Фрагмент молекулы ДНК, включающий промотор, транскрибируемую последовательность и терминатор образует единицу транскрипции – транцкриптон.
В процессе синтеза по мере продвижения РНК-полимеразы вдоль молекулы ДНК, пройденные ею одноцепочечные участки ДНК вновь объединяются в двойную спираль. Образуемая в ходе транскрипции мРНК содержит точную копию информации, записанной в соответствующем участке ДНК. Тройки рядом стоящих нуклеотидов мРНК, шифрующие аминокислоты, называют кодонами. Последовательность кодонов мРНК шифрует последовательность аминокислот в пептидной цепи. Кодонам мРНК соотвествуют определенные аминокислоты.
5. Кроме митотического деления, у одноклеточных обнаружен также половой процесс, который заключается обычно в слиянии двух половых клеток - гамет. Формы полового процесса у одноклеточных организмов можно объединить в две группы: конъюгацию, при которой специальные половые клетки не образуются, и гаметическую копуляцию, когда формируются половые элементы и происходит их попарное слияние.
Билет
1. Клетка представляет собой основную единицу строения всех живых организмов, клетки животных и растений сходны по строению. Изучение химической организации клетки привело к выводу, что именно химические процессы лежат в основе ее жизни, что клетки всех организмов сходны по химическому составу, у них однотипно протекают основные процессы обмена веществ. Современная клеточная теория включает следующие положения:
1.Клетка является структурно-функциональной единицей, а также единицей развития всех живых организмов;
2.Клеткам присуще мембранное строение;
3.Ядро - главная составляющая клетки;
4.размножение клеток происходит путем их деления, и каждая новая клетка образуется в результате деления исходной (материнской) клетки;
Клеточное строение организмов - доказательство единого происхождения растений и животных;
Изучение клеток разнообразных одноклеточных и многоклеточных организмов показало, что по своему строению они разделяются на две группы.. 1.Организмы имеющие наиболее простое строение клеток называются доядерными (прокариотами), так как у них нет оформленного ядра и нет многих структур, которые называют органоидами.
2.Другую группу называют ядерными (эукариотическими). Эти клетки имеют ядро и органоиды, выполняющие специфические функции.
2. В Немембранные органоиды цитоплазмы включают фибриллярные структуры и рибосомы. К фибриллярным структурам относятся микрофиламенты, микрофибриллы и микротрубочки. Микрофиламенты — тонкие нити сократительных белков.Могут распологаться в разных участках цитоплазмы Они образуют пучки или рыхлую сеть. Микрофиламенты могут выполнять и каркасную функцию. Микрофибриллы, или промежуточные фибриллы образуют пучки и располагаются в центральной части вокруг ядра. В различных типах клеток микрофибриллы построены из разных белков — способных к сополимеризации; эти структуры выполняют опорно-скелетную функцию. Микротрубочки — полые длинные неветвящиеся цилиндры.Стенка образована 13-ю нитями скрученными по спирали одна над другой. В митозе образуют веретено деления, они входят в состав постоянных органоидов —-центриолей,ресничек и жгутиков. Микротрубочки состоят из белков тубулинов. Они являются динамичными структурами, способными к сборке и разборке, выполняют опорно-скелетную функцию и участвуют в двигательных реакциях клетки.
Помимо органоидов, в цитоплазме некоторых типов клеток имеются непостоянные компоненты — включения. Они представляют собой особую форму углеводов и липидов в виде жировых капель.
3.Клеточный цикл -это период существования клетки от момента ее образования путем деления материнской клетки до собственного деления или смерти. Клеточный цикл составляют два периода:
1) период клеточного роста-интерфаза. идет синтез ДНК и белков и осуществляется подготовка к делению клетки.
Интерфаза состоит из нескольких периодов: G1-фазы- фазы начального роста, во время которой идет синтез мРНК, белков, других клеточных компонентов; S-фазы (синтетическая), во время которой идет репликация ДНК клеточного ядра, также происходит удвоение центриолей (если они есть). G2-фазы, во время которой идет подготовка к митозу.У дифференцировавшихся клеток, которые более не делятся, в клеточном цикле может отсутствовать G1 фаза. Такие клетки находятся в фазе покоя G0.
2) период клеточного деления-митоз.
Период клеточного деления (фаза М) включает две стадии:
кариокинез (деление клеточного ядра);
цитокинез (деление цитоплазмы).
4.ДНК представляет собой спираль, состоящую из двух полинуклеотидных цепей.В состав нуклеотидов ДНК входят: азотистое основание, дезоксирибоза и остаток фосфорной кислоты. Азотистые основания делят на пуриновые (аденин и гуанин) и пиримидиновые (тимин и цитозин). Две цепи нуклеотидов соединяются между собой через азотистые основания по принципу комплементарности: между аденином и тимином возникают две водородные связи, между гуанином и цитозином — три.
Функции ДНК:1) обеспечивает сохранение и передачу генетической информации от клетки к клетке и от организма к организму, что связано с ее способностью к репликации;
2) регуляция всех процессов, происходящих в клетке, обеспечиваемая способностью к транскрипции с последующей трансляцией.
Процесс самовоспроизведения ДНК называется репликацией. Репликация обеспечивает копирование генетической информации и передачу ее из поколения в поколение, генетическую идентичность дочерних клеток, образующихся в результате митоза, и постоянство числа хромосом при митотическом делении клетки.
5. Размножение –одно из основных свойств живого.Размножение подразумевает способность организмов производить себе подобных особей.известны две основные формы размножения половое и бесполое.
Половой процесс. Половое размножение отличается наличием полового процесса, который обеспечивает обмен наследственной информацией и создает условия для возникновения наследственной изменчивости. В нем, как правило, участвуют две особи — женская и мужская. В результате оплодотворения, т. е. слияния женской и мужской гамет, образуется диплоидная зигота с новой комбинацией наследственных признаков. Половое размножение по сравнению с бесполым обеспечивает появление наследственно более разнообразного потомства. Формами полового процесса являются конъюгация и копуляция.
1.Конъюгация — своеобразная форма полового процесса, при которой оплодотворение происходит путем взаимного обмена мигрирующими ядрами, перемещающимися из одной клетки в другую по цитоплазматическому мостику, образуемому двумя особями. При конъюгации обычно не происходит увеличения количества особей, но происходит обмен генетическим материалом между клетками, что обеспечивает перекомбинацию наследственных свойств. Конъюгация типична для ресничных простейших (например, инфузорий), некоторых водорослей (спирогиры).2.Копуляция (гаметогамия) — форма полового процесса, при которой две различающиеся по полу клетки — гаметы — сливаются и образуют зиготу. При этом ядра гамет образуют одно ядро зиготы.
При бесполом размножении потомки развиваются из одной материнской клетки или группы клеток. Выделяют несколько форм бесполого размножения
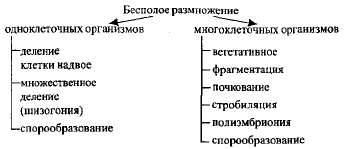
Билет.
1.Биология -совокупность наук о живой природе. Предмет изучения Биология — все проявления жизни: строение и функции живых существ и их природных сообществ, их распространение, происхождение и развитие, связи друг с другом и с неживой природой. Задачи Биология состоят в изучении всех биологических закономерностей, раскрытии сущности жизни и её проявлений с целью познания и управления ими. Термин «Биология» предложен в 1802 независимо друг от друга двумя учёными — французом Ж. Биология Ламарком и немцем Г. Р. Тревиранусом.
Система биологических наук чрезвычайно многопланова, что обусловлено как многообразием проявлений жизни, так и разнообразием форм, методов и целей исследования живых объектов, изучением живого на разных уровнях его организации. Одними из первых в Биология сложились науки о животных — зоология и растениях — ботаника, а также анатомия и физиология человека — основа медицины. Другие крупные разделы Биология, выделяемые по объектам исследования, — микробиология — наука о микроорганизмах, гидробиология — наука об организмах, населяющих водную среду, и т.д. Внутри Биологии сформировались более узкие дисциплины; в пределах зоологии — изучающие млекопитающих — териология, птиц — орнитология, пресмыкающихся и земноводных — герпетология, рыб и рыбообразных — ихтиология, насекомых — энтомология, клещей — акарология, моллюсков — малакология, простейших — протозоология; внутри ботаники — изучающие водоросли — альгология, грибы — микология, лишайники — лихенология, мхи — бриология, деревья и кустарники — дендрология и т.д. Подразделение дисциплин иногда идёт ещё глубже. Многообразие организмов и распределение их по группам изучают систематика животных и систематика растений. Биологию можно подразделить на неонтологию, изучающую современный органический мир, и палеонтологию — науку о вымерших животных (палеозоология) и растениях (палеоботаника).
Другой аспект классификации биологических дисциплин — по исследуемым свойствам и проявлениям живого. Форму и строение организмов изучают морфологические дисциплины; образ жизни животных и растений и их взаимоотношения с условиями внешней среды — экология; изучение разных функций живых существ — область исследований физиологии животных и физиологии растений; предмет исследований генетики — закономерности наследственности и изменчивости; этологии — закономерности поведения животных; закономерности индивидуального развития изучает эмбриология или в более широком современном понимании — биология развития; закономерности исторического развития — эволюционное учение. Каждая из названных дисциплин делится на ряд более частных (например, морфология — на функциональную, сравнительную и др.). Одновременно происходит взаимопроникновение и слияние разных отраслей Биология с образованием сложных сочетаний, например гисто-, цито- или эмбриофизиология, цитогенетика, эволюционная и экологическая генетика и др. Анатомия изучает строение органов и их систем макроскопически; микроструктуру тканей изучает гистология, клеток — цитология, а строение клеточного ядра — кариология. В то же время и гистология, и цитология, и кариология исследуют не только строение соответствующих структур, но и их функции и биохимические свойства.
2. Органоиды специального назначения - (имеются только в клетках высокоспециализированных тканей и обеспечивают выполнение строгоспецифических функций этих тканей): в эпителиальных клетках - реснички, микроворсинки, тонофибриллы; в нейральных тканях - нейрофибриллы и базофильное вещество; в мышечных тканях - миофибриллы. У многих растительных и животных клеток имеются органоиды специального назначения: реснички, выполняющие функцию движения (инфузории, клетки дыхательных путей), жгутики (простейшие одноклеточные, мужские половые клетки у животных и растений и др.). Жгутики и реснички — органоиды нитевидной формы, представляют собой аксонему, ограниченную мембраной. Аксонема — цилиндрическая структура; стенка цилиндра образована девятью парами микротрубочек, в его центре находятся две одиночные микротрубочки. В основании аксонемы находятся базальные тельца, представленные двумя взаимно перпендикулярными центриолями (каждое базальное тельце состоит из девяти триплетов микротрубочек, в его центре микротрубочек нет). Миофибриллы состоят из актиновых и миозиновых миофиламентов, обеспечивающих сокращение мышечных клеток.
3. Хромосомы в клетке под микроскопом можно увидеть только во время деления (митоза, во время стадии - метафазы), такие хромосомы называются - метафазные. Когда клетка не делится хромосомы имеют вид тонких, темноокрашенных нитей, называемых хроматином.
Хроматин представляет собой дезоксирибонуклеопротеид (ДНП. Метафазная хромосома состоит из двух продольных нитей ДНП - хроматид, соединенных друг с другом в области первичной перетяжки. Центромера делит тело хромосомы на два плеча. В зависимости от расположения первичной перетяжки различают следующие типы хромосом: равноплечие (метацентрические), когда центромера расположена посередине, а плечи примерон равной длины; неравноплечие (субметацентрические), когда центромера смещена от середины хромосомы, а плечи неравной длины; палочковидные (акроцентрические), когда центромера смещена к одному концу хромосомы и одно плечо очень короткое
Для процедуры определения кариотипа могут быть использованы любые популяции делящихся клеток, для определения человеческого кариотипа используется либо одноядерные лейкоциты, извлечённые из пробы крови, деление которых провоцируется добавлением митогенов, либо культуры клеток, интенсивно делящихся в норме (фибробласты кожи, клетки костного мозга). Обогащение популяции клеточной культуры производится остановкой деления клеток на стадии метафазы митоза добавлением колхицина — алкалоида, блокирующего образование микротрубочек и «растягивание» хромосом к полюсам деления клетки и препятствующего тем самым завершению митоза.
Полученные клетки в стадии метафазы фиксируются, окрашиваются и фотографируются под микроскопом; из набора получившихся фотографий формируются т. н. систематизированный кариотип — нумерованный набор пар гомологичных хромосом (аутосом), изображения хромосом при этом ориентируются вертикально короткими плечами вверх, их нумерация производится в порядке убывания размеров, пара половых хромосом помещается в конец набора.
4. Открытия экзон-интронной организации эукариотических генов и возможности альтернативного сплайсинга показали, что одна и та же нуклеотидная последовательность первичного транскрипта может обеспечить синтез нескольких полипептидных цепей с разными функциями или их модифицированных аналогов. Например, в митохондриях дрожжей имеется ген box (или cob), кодирующий дыхательный фермент цитохром b. Он может существовать в двух формах. «Длинный» ген, состоящий из 6400 п. н., имеет 6 экзонов общей протяженностью 1155 п.н. и 5 интронов. Короткая форма гена состоит из 3300 п.н. и имеет 2 интрона. Она фактически представляет собой лишенный первых трех интронов «длинный» ген. Обе формы гена одинаково хорошо экспрессируются.
После удаления первого интрона «длинного» гена box на основе объединенной нуклеотидной последовательности двух первых экзонов и части нуклеотидов второго интрона образуется матрица для самостоятельного белка — РНК-матуразы. Функцией РНК-матуразы является обеспечение следующего этапа сплайсинга — удаление второго интрона из первичного транскрипта и в конечном счете образование матрицы для цитохрома b.
У вирусов и бактерий описана ситуация, когда один ген может одновременно являться частью другого гена или некоторая нуклеотидная последовательность ДНК может быть составной частью двух разных перекрывающихся генов. Например, на физической карте генома фага ФХ174 видно, что последовательность гена В располагается внутри гена А, а ген Е является частью последовательности гена D. Этой особенностью организации генома фага удалось объяснить существующее несоответствие между относительно небольшим его размером (он состоит из 5386 нуклеотидов) и числом аминокислотных остатков во всех синтезируемых белках, которое превышает теоретически допустимое при данной емкости генома. Возможность сборки разных пептидных цепей на мРНК, синтезированной с перекрывающихся генов (А и В или Е и D), обеспечивается наличием внутри этой мРНК участков связывания с рибосомами. Это позволяет начать трансляцию другого пептида с новой точки отсчета.
Центральная догма молекулярной биологии — обобщающее наблюдаемое в природе правило реализации генетической информации: информация передаётся от нуклеиновых кислот к белку, но не в обратном направлении. Правило было сформулировано Френсисом Криком в 1958 году и приведено в соответствие с накопившимися к тому времени данными в 1970 году. Переход генетической информации от ДНК к РНК и от РНК к белку является универсальным для всех без исключения клеточных организмов, лежит в основе биосинтеза макромолекул. Репликации генома соответствует информационный переход ДНК → ДНК. В природе встречаются также переходы РНК → РНК и РНК → ДНК (например у некоторых вирусов), а также изменение конформации белков, передаваемое от молекулы к молекуле.
5. в процессе мейоза образуются половые клетки, с гаплоидным набором хромосом, достигается благодаря однократной редупликации ДНК, для двух последовательных делений мейоза, а также благодаря образованию в начале первого мейотического деления пар гомологичных хромосом и дальнейшего их расхождения в дочерние клетки.
Date: 2015-04-23; view: 890; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |