Полезное:
Как сделать разговор полезным и приятным
Как сделать объемную звезду своими руками
Как сделать то, что делать не хочется?
Как сделать погремушку
Как сделать так чтобы женщины сами знакомились с вами
Как сделать идею коммерческой
Как сделать хорошую растяжку ног?
Как сделать наш разум здоровым?
Как сделать, чтобы люди обманывали меньше
Вопрос 4. Как сделать так, чтобы вас уважали и ценили?
Как сделать лучше себе и другим людям
Как сделать свидание интересным?

Категории:
АрхитектураАстрономияБиологияГеографияГеологияИнформатикаИскусствоИсторияКулинарияКультураМаркетингМатематикаМедицинаМенеджментОхрана трудаПравоПроизводствоПсихологияРелигияСоциологияСпортТехникаФизикаФилософияХимияЭкологияЭкономикаЭлектроника
Возникновение биосферы
Нефть и все другие горючие полезные ископаемые, так же как рассеянное органическое вещество осадочных пород, генетически связаны с живым веществом нашей планеты, с биосферой прошлых геологических эпох. Проблема происхождения нефти, нижний возрастной предел ее образования тесно связаны с возрастом возникновения жизни на Земле. Согласно наиболее распространенной гипотезе, Земля возникла 4,8-5 млрд лет назад в результате слипания первичного вещества холодных тел — плане-тознмалей, затем произошел ее разогрев вследствие повышенной теплогенерации. Источники энергии — радиоактивный распад, импактные воздействия, ультрафиолетовое излучение, сейсмичность, приливные возмущения и др. В результате произошла дифференциация вещества первичной Земли и сформировались ядро, мантия и земная кора, близкая по составу к современной. Дифференциация вещества вызвала выделение газов и формирование первичных океанов и атмосферы. Первичная атмосфера от-личачась от современной. Она имела восстановительный характер, в ее составе были гелий и водород, которые быстро улетучились, метан, пары воды, аммиак, СО, СО2. Свободный кислород отс\тствовал. За счет высокой активности этих веществ, очевидно, образовывались полимеры, содержащие С, N, О и другие онофильные элементы, т.е. первые органические вещества возникали путем абиогенного синтеза.
Идея о том, что современные биологические молекулы могли в прошлом возникнуть абиогенным путем, была высказана впервые А.И. Опариным, затем Дж. Холдейном еще в середине 20-х годов XX в. Экспериментальные работы в этом направлении началась только в 50-х годах в разных лабораториях. Наиболее сенсационный результат был получен в лаборатории Чикагского университета Стэнли Миллером, который в результате облучения УФ-лучами газовой смеси, содержащий метан, аммиак, водород и воду, а также использованием электрического разряда синтезировал мочевину, муравьиную кислоту и формальдегид. Затем были
золучены аминокислоты, альдегиды, аденин, глицин и другие органические соединения. К настоящему времени абиогенным путем синтезирован весь ряд аминокислот, входящих в белки.
В чем же разница живого и неживого вещества? Синтезиро-юнные в лабораториях, видимо, как и существующие на первич-яой Земле сгустки белка, не могли воспроизводить себе подобных. Для этого необходима «матрица», каковой в биологических клетках и являются ДНК и РНК, т.е. основные молекулы жизни — биополимеры: белки (полипептиды) и ДНК и РНК (поли-нуклеиды). Основной признак жизни — самовоспроизведение, самообновление белковых тел, в основе которых лежит самореп-дикация или удвоение молекулы ДНК с передачей рождающейся клетке генетической информации. Академик В.И. Гольданский, определяя жизнь как «форму существования биополимерных тел iсистем), способных к саморепликации в условиях постоянного обмена веществ и энергией с окружающей средой», подчеркивал, что это не единственная особенность, отличающая биоорганический мир от неживой природы.
Второе важное отличие живой природы состоит в хиральной чистоте биополимеров (Гольданский, 1986). В чем же суть этого отличия? Хиральность (от греч: xeip — рука) — способность молекул образовывать зеркально-антиподные изомеры, которые называют оптическими изомерами. Они отличаются друг от друга тем, что вращают плоскость поляризованного света в противоположные стороны: в правую D-изомеры — правые, в левую — L-изо-меры — левые. Живой природе присуща практически абсолютная хиральная чистота: белки содержат только левые аминокислоты, а нуклеиновые кислоты — только правые сахара. В исходном предбиологическом органическом веществе зеркальных антиподов не существовало, т.е. «правые» и «левые» изомеры присутствовали в равных количествах. Такие смеси называются рацемическими. Это подтверждено многочисленными экспериментами по синтезу органических веществ, проводимых более 40 лет в разных лабораториях мира. В результате каких сил и процессов была разрушена оптическая симметрия органических молекул — это до сих пор нерешенный вопрос для биохимиков, химиков, биологов и геохимиков.
На вопрос «когда появилась жизнь?» очевиден ответ — когда появилась ДНК, и РНК внедрилась в структуру синтезированного белка. Свидетельства существования биологической жизни в породах раннепротерозойского и архейского возраста получены в разных районах мира — шунгиты Карелии, метаморфические сланцы и метакварциты Гренландии, Южной Африки. Все они содержат Сорг; при помощи электронной микроскопии в них обнаружены форменные остатки, методом хроматомасс-спектромет-рии идентифицированы хемофоссилии.
К числу наиболее древних определений биогенного ОВ относится формация Фиг-Три в Южной Африке, возраст которой 3,1 млрд лет. В кремнистых сланцах формации обнаружены форменные остатки, идентифицированные как «окаменелости бактерий». Хроматограмма экстракта из этих пород имеет нормальное распределение н-алканов С14-С25, в малых количествах присутствуют пристан и фитан (Кальвин, 1971). По-другому выглядит хроматограмма экстракта из более древних пород формации Онфервахт (возраст 3,7 млрд лет). На ней на фоне гладкой кривой выделяются отдельные пики, обычно свойственные н-алка-нам биогенного генезиса, т.е. эта хроматограмма идентифицируется как смесь абиогенных и биогенных УВ. М. Кальвин сделал это заключение с большой осторожностью, отдавая себе отчет в том, что для такого вывода (граница жизнь-преджизнь) требуются очень веские доказательства. На рис. 3.1 приведена данная М. Кальвиным геохронологическая таблица химической и биологической эволюции, на которой четкая граница между этими двумя важнейшими этапами в эволюции Земли отсутствует, она лежит в интервалах 3,1-3,7 млрд лет.
Анализ всех данных палеогеохимии, палеобиологии и геохимии, проведенный Дж. У. Шопфом, позволяет предположить, что жизнь появилась на Земле более 3,5 млрд лет назад и, вероятно, ранее этого рубежа в органическом мире уже были развиты такие явления, как анаэробная хемогетеротрофия и анаэробная фитоав-тотрофия (Соколов, Федонкин, 1988).
Считают, что первые организмы были анаэробные гетеротро-фы — бродильщики; они потребляли органические вещества, синтезированные абиогенным путем. Когда запас этих веществ был исчерпан, появились первые автотрофы. Они черпали энергию для жизни, расщепляя H2S, выделяя при этом серу, — окси-генный фотосинтез, или анаэробный хемосинтез; затем они научились потреблять водород, расщепляя воду и выделяя кислород — оксигенный фотосинтез. Формирование ОВ протекало согласно реакциям:
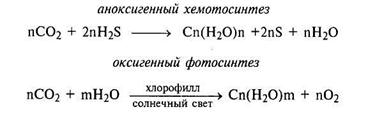
Выделение кислорода в атмосферу и накопление его привело к формированию озонового слоя, благодаря которому жизнь смогла выйти на сушу.
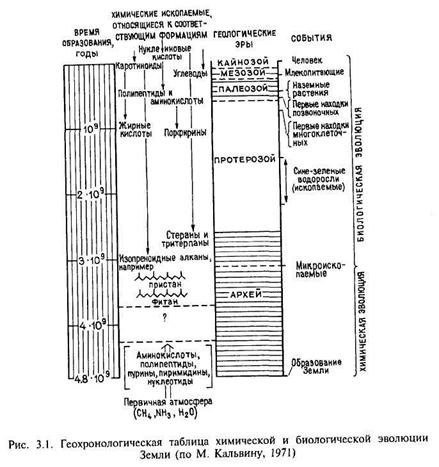
Первые организмы (архебактерии, бактерии, цианобактерии) были прокариотами, т.е. у них отсутствовало ядро и половое деление, ДНК сосредоточена в центре клетки.
Существуют разные точки зрения, когда появился кислород в атмосфере. Согласно одной из них, накопление О2 в атмосфере шло медленно и плавно, достигнув современного уровня в течение фанерозоя, так как прокариотная система не могла обеспечить более 1% свободного кислорода в атмосфере. Более распространена идея, что свободный кислород начал играть заметную роль в атмосфере начиная с 1,8-2,1 млрд лет назад — именно к этому времени приурочено появление красноцветных формаций, широко распространенных практически на всех континентах. Существует также гипотеза, основанная на сходстве изотопного состава серы фанерозойских и архейских образований, что аэробная фотоавтотрофия существовала уже 2,9-3 млрд лет назад.
Эукариоты появились 1,5 млрд лет назад, они имели клетку с четко выраженным ядром; первые эукариоты были однополыми. В интервале 0,8-1,0 млрд лет появились разнополые эукариоты, которые дали толчок бурному развитию разных форм жизни; к концу протерозоя появились зеленые, бурые, красные водоросли. На границе кембрия-докембрия появление многоклеточных организмов повлекло за собой бурное развитие жизни — фауны и флоры. Возможно, что бесскелетная фауна появилась раньше. В вендских отложениях (600 млн лет) найдены отпечатки крупных бесскелетных организмов (эдиакарская фауна), но точная принадлежность этой группы ископаемых не определена. Первые наземные растения фиксируются в конце силура, расцвет их начинается с карбона и продолжается с постоянным увеличением видов вплоть до кайнозоя, с начала кайнозоя — млекопитающие, а в конце кайнозоя — человек (Homo sapiens) — вершина эволюции живого вещества.
Основные биопродуценты и их эволюция
Несмотря на единство элементного и компонентного составов различных представителей живого вещества, в их химическом составе и молекулярной структуре наблюдаются заметные отличия.
Живое вещество любого участка суши или моря характеризуется определенными биоценозами. Биоценоз — это совокупность всех живых организмов (животных, растений, грибов и др.), совместно населяющих какой-либо участок суши и (или) водоема. В любом биоценозе по типу питания выделяются три группы организмов: продуценты, консументы и редуценты. Продуценты — живые организмы, производящие живое вещество из неживого, автотрофы, например зеленые растения, водоросли; консументы — потребители, питающиеся ОВ продуцентов, — гетеротро-фы — все животные; редуценты — грибы и бактерии, разлагающие ОВ как консументов, так и продуцентов, минерализующие его.
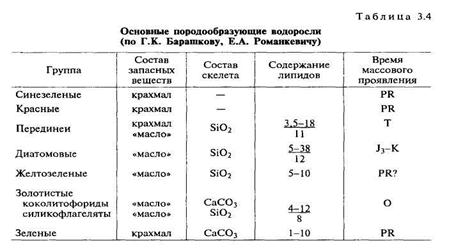
Основным продуцентом ОВ в современных водоемах является фитопланктон. Поскольку НМ потенциал ОВ в основном определяется долей липидных и липоидных компонентов в исходном ОВ, то именно распределение этой группы соединений в основных продуцентах представляет особый интерес (см. табл. 3.2). В табл. 3.4 приводится содержание липидов в разных группах водорослей. Повышенной концентрацией липидов отличаются диатомовые водоросли, по биопродуктивности первое место занимают так же диатомовые, затем следуют динофлагелла-ты, относящиеся к группе перидиней. Современные диатомовые водоросли синтезируют более 50% ОВ. Широкое распростране-
ние они получили с конца юрского периода, хотя некоторые исследователи считают, что первые диатомеи появились уже к началу палеозоя. Диатомеи содержат повышенные концентрации внутриклеточных липидов (до 38% сухого веса); кроме того, ли-пиды входят в состав клеточных мембран и запасных систем. Состав липидов непостоянен, соотношение свободных и связанных жирных кислот, их распределение различны. Так, у диатомеи Sceletonema costatum в общем количестве липоидов жиры составляют 34%, свободные жирные кислоты — 15%, глицериды — 11%, галактолипиды — 12%. Преобладают жирные кислоты с не-разветвленной цепью и четным числом атомов С, причем незначительно превалируют ненасыщенные кислоты: C16 — 70%, С14 — 10%, С20 — 5%, очень мало C18 и С22 (Барашков, 1972).
Характерной особенностью диатомовых и некоторых других водорослей (золотистых, желтозеленых, перидиней) является их способность накапливать в виде запасных веществ липиды, причем наиболее нейтральные. Благодаря отложению этого легкого резервного вещества диатомеи имеют достаточную подвижность, несмотря на наличие тяжелой кремниевой оболочки. Содержание и состав запасных липидов зависят от разных экологических факторов: освещенности, содержания минеральных веществ. Установлено, что при недостатке азота, при голодании в диатоме-ях резко возрастает доля липидов в ОВ. Дополнительные накопления липидов происходят в результате стрессовых ситуаций: облучение, холод, тепло, нарушение солевого обмена, воздействие химических стимуляторов. В эксперементальных условиях доля липидов возрастала до 70%. Важнейшим компонентом запасных
липидов являются длинноцепочные жирные кислоты, в основном ненасыщенные.
Накопив запасные вещества, водоросль может их сама израсходовать в процессе жизнедеятельности. Для того чтобы запасные липиды попали в осадок, необходимо отмирание организма в момент накопления этих резервных веществ. Таким образом, на состав ОВ осадка влияют не столько экологические условия жизни основных биопродуцентов, сколько условия их отмирания, т.е танатоценозы.
Следующая по численности группа альгофитопланктона представлена динофлагеллатами, относящимися к группе периденей. или панцирных жгутиконосцев. Перидинеи в массовых количествах встречаются в основном в теплых морях. В их клетках несколько повышено содержание липидов — от 3 до 18%, в среднем 11%. В составе жирных кислот присутствуют С18, С20, С22. резко преобладает C18 (до 57%). Установлены также стероиды. Характерный биомаркер — 4-метилстероид, обнаруженный в повышенных концентрациях в ОВ рэтских горючих сланцев Западной Англии, рассматривается как показатель исходной биомассы динофлагеллат. Расцвет перидинеи был в поздней юре и мелу, в кайнозое их распространение пошло на спад, но и в современных водоемах, морских и пресноводных, они еще достаточно широко развиты. Единичные их формы описаны в силуре в Черногории, но широкое распространение типичные перидинеи получили только с юрского периода.
Золотистые водоросли более распространены среди ископаемых форм, окраска их обусловлена наличием желтого фермента — финохризина. Примитивные формы золотистых водорослей известны с кембрия, более сложные — с мела. Наиболее распространены два подкласса: коколитофориды (coccolitophorea) — очень мелкие нанопланктонные формы, обитающие в теплых морях и образующие известковый ил; силикофлагеллаты (silicoflu-gellaiae) — водоросли с кремниевой функцией, кремниевые жгутиковые. Эти водоросли достигли широкого распространения с сенона, встречаются они и в отложениях юры, имеются сведения о нахождении примитивных форм в ордовике. Это очень мелкие, исключительно морские формы, обитают в основном в области литорали. В биохимическом составе этих водорослей изучены только жирные кислоты; их состав: C18 — 16—27%,С14 — 11%, С16 — 7-10%. Вкачестве резервных веществ современные формы накапливают липиды.
Жвлтозеленые водоросли значительно менее распространены; они играют важную роль в формировании озерных отложений, обогащенных ОВ. Свое название эта группа водорослей получила благодаря содержанию наряду с хлорофиллом желтых пигментов — ксантофилла и каротина. Желтозеленые водоросли обнару-
хивают большое сходство с зелеными водорослями. Особый интерес представляет водоросль Batryococcus, широко распространенная в современных сапропелевых илах и ископаемых сапро-зелитах, в горючих сланцах, как озерных, так и морских. Так, содержание УВ в варпалотских горючих сланцах Венгрии прямо пропорционально количеству остатков водорослей Batryococcus. Батриококкус является породообразующей водорослью так называемых торбанитов — озерных горючих сланцев Австралии пермского возраста.
Способность накапливать этими водорослями в особых усло-зиях липиды в больших количествах может быть использована для получения синтетической нефти. Ученые Иерусалима в лаборатории получили углеводороды из водорослей Batryococcus, выращенных в теплой соленой воде. В США были выполнены подобные эксперименты: в воду, населенную водорослями Batryococcus, добавляли азот, микроэлементы и продували диоксид углерода, при этом резко возрастала биопродуктивность водорослей и содержание в них липидов. Квадратный метр поверхности такого водоема ежедневно может производить 50 г ОВ, из них 70% составляют липиды, 80% которых используется для получения дизельного топлива.
Наиболее широкое стратиграфическое распространение свойственно синезеленым водорослям. Они относятся к прокариотам, что сближает их с бактериями. Есть и другие признаки, более свойственные бактериям: строение клеточной стенки, наличие газовых вакуолей, способность к фиксации азота и др. В настоящее время их чаще называют цианобактериями. Они существуют на Земле более 3 млрд лет. Автотрофные формы при фотосинтезе используют СО2 и выделяют кислород; благодаря их жизнедеятельности была создана кислородная атмосфера Земли. В течение всей истории своего развития они не претерпели изменений. В протерозойских бассейнах они были подавляющей формой жизни и поставщиком ОВ. Многими исследователями отмечались консервативность цианобактерий, их экологическая выносливость. Синезеленый цвет определяется наличием синего и бурого пигментов в сочетании с хлорофиллом. Некоторые формы имеют и другие пигменты - от красного до черного. Эти водоросли токсичны, хищны, подавляют развитие других водорослей и зоопланктона, радиорезистентны, приспособлены жить в темноте, в горячих и холодных водах. Очень важным свойством этих водорослей является антибактериальное действие их липидов (циано-фитина и хлороллина). Это предопределило устойчивость ОВ си-незеленых (как и некоторых зеленых водорослей) к микробному разрушению. Цианобактерий представлены как одноклеточными, так и многоклеточными формами.
В цианобактериях обнаружены чрезвычайно устойчивые биополимеры, но существенно иного типа, чем в батриококкусах. Эти биополимеры образуются в оболочке клеток живого вещества и характеризуются аморфной структурой, благодаря им формируется аморфный кероген. Такие структуры керогена встречены как в современных осадках, так и в страмотолитах докембрийского возраста. Благодаря этому ОВ синезеленых водорослей имеет достаточно постоянный и устойчивый состав, оно несколько обеднено гетероэлементами и обогащено водородом. Несмотря на невысокое в среднем содержание липидных компонентов (до 12%), оно характеризуется высоким нефтематеринским потенциалом.
Основную массу ОВ поставляют продуценты — основа пищевой цепи. Следующая группа пищевой цепи — копеподы, крупнейшая группа зоопланктона. Копеподы питаются непосредственно фитопланктоном, видимо, поэтому существует сходство между составом липидной фракции копепод и фитопланктона. Изопреновый углеводород — пристан, присутствующий во многих современных осадках, видимо, имеет тот же источник — он является основным компонентом жировых телец копепод из отряда Calonoida.
При переходе по пищевой цепи от продуцента к следующему звену пищевой цепи масса ОВ сокращается на порядок. В ряду фитопланктон → копеподы → рыбы → хищники масса ОВ сократилась в 1000 раз. Несмотря на это отдельные группы зоопланктона, видимо, вносят заметный вклад в ОВ и (или) влияют на его исходный состав. Так, ОВ силурийских граптолитовых сланцев характеризуется повышенными содержаниями азота за счет зоопланктона.
Вклад бактериального ОВ в исходное ОВ неоднозначно оценивается различными исследователями. Ведущая роль бактерий в преобразовании ОВ, в формировании окислительно-восстановительной обстановки, в воздействии на минеральную составляющую породы признается всеми. Бактерии — наиболее распространенные представители живого вещества. Благодаря им зона действия живого вещества расширяется до глубины в несколько километров, охватывает высокотемпературные (более 100°С) зоны, такие как области развития «курильщиков», горячие озера. На основании такой «повсюдности» бактерий было предложено выделять сферу их жизнедеятельности как бактериосферу (Соколов, 1993).
Бактерии — мельчайшие организмы (ультрапланктон), размер которых не превышает 5 мкм. Бактерии являются автотрофами, гетеротрофами и олиготрофами. Автотрофные бактерии включают фотосинтезирующие формы, использующие для синтеза ОВ из минеральных веществ солнечную энергию, и хемосинтезирую-щие, использующие для этой цели химическую энергию. Авто-
трофные бактерии — это нитрофикаторы, сульфатредуцирующие, железобактерии и др.
Большая часть бактерий — гетеротрофы, они существуют за счет органических веществ других организмов, за счет разложения сложных органических молекул. Гетеротрофы, использующие для питания мертвые ОВ (некрому), называются сапрофитами, или гнилостными; те же бактерии, которые живут в теле живых организмов, называются паразитами.
По отношению к кислороду бактерии подразделяются на аэробные, развивающиеся в присутствии кислорода, анаэробные — при отсутствии свободного кислорода, факультативно-анаэробные, растущие в тех и других условиях. Бактерии, развивающиеся в высокоминерализованных средах (более 100 г/л), называются галофильными. Анаэробные гетеротрофы — это метанобразующие, метанокисляюшие, молочнокислые, маслянокислые, гнилостные бактерии и др.
Характерной чертой бактерий является автолиз — самоуничтожение своей некромы. Автолизу препятствует сорбция бактериального ОВ минеральными частицами и резко восстановительная обстановка. Возможно, что автолиз уничтожает не все ОВ бактерии, а только наиболее подверженные микробиальному окислению компоненты, липидная же составляющая или часть ее сохраняется.
Согласно расчетам Н.В. Лопатина, доля некромы бактерий в составе ОВ к концу диагенеза может составлять ориентировочно в илах озер до 25-35%, в мелководных морских заливах и лагунах 10-15%, в шельфовой зоне океана 5-10%, в океанических осадках менее 0,5% (Лопатин, 1982).
Таким образом, геохимическая роль бактерий заключается прежде всего в разложении огромных масс растительных и животных остатков. Велика роль их в круговороте С, N, P, S, Fe, Мn и др. С жизнедеятельностью бактерий связаны процессы биодеградации нефти и нафтидов, образования и разрушения серы и сульфидных руд, окисления метана в угольных месторождениях и т.д. Если вопрос о вкладе бактериального ОВ в исходное ОВ до сих пор дисскусионен, то участие липидной составляющей бактерий в формировании состава ОВ бесспорно.
Бактерии имеют изменчивый состав: 80% и более составляет вода, 20% — ОВ. По современным оценкам, 30-40% мертвого ОВ, потребляемого бактериями, идет на построение живого вещества самой бактерии, остальные 60-70% служат для получения энергии и после изменений превращаются в минеральные осадки. Их химический состав: С — 50%, О — 20%, Н — 8%, N — 5-10%, S — 1%, в малых количествах присутствуют магний, кальций, марганец, медь, цинк. Групповой состав: белки — 50-60%,
материал клеточных мембран — 20%, на 20-40% состоящих из липидов, 10% липиды, липоиды, 10% ДНК, РНК. Жирные кислоты, извлекаемые из бактерий, обычно представлены соединениями ряда С10-С20, преобладают кислоты с разветвленной цепью: изо- и антеизоконфигурациями. Помимо жиров многие бактерии способны накапливать полифосфаты, полисахариды и серу. В составе ОВ бактерий отмечены также тритерпаны ряда гопана, в частности бисноргопаны, гаммацераны.
Высшие растения также являются поставщиком органического вещества в исходное ОВ аквальных осадков. ОВ высших растений представлено главным образом целлюлозой и лигнином, доля липоидных компонентов в них в целом мала. Однако заметное количество липидов и липоидов накапливается в листьях, спорах, пыльце, коре, плодах и семенах. Липиды высших растений характеризуются некоторой специфичностью, например, среди н-алканов от С10 до С40 отмечается повышенная концентрация УВ высокомолекулярных — С27-С29, а также значительное (до 10 раз) преобладание нечетных УВ над четными. В строении нерастворимой части керогена участвуют компоненты углеводов, белков и лигнина.
Состав и тип захороняющегося в осадках ОВ определяется прежде всего соотношением разных групп организмов-поставщиков, фациальной и физико-химической обстановками осадкона-копления. Абстрагируясь от условий осадконакопления, проследим, как химический состав ОВ осадка в значительной степени изменялся в зависимости от эволюционного уровня развития групп организмов-поставщиков ОВ.
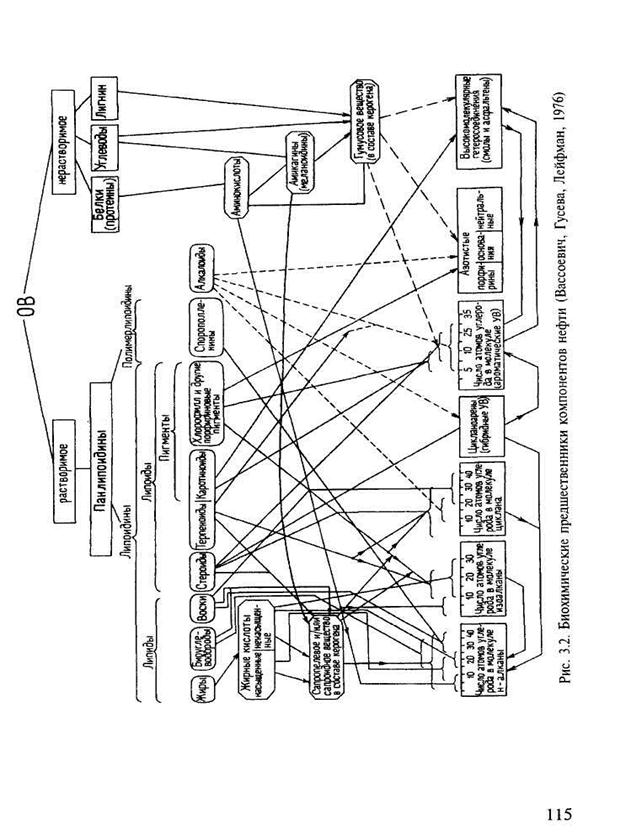
Биохимические предшественники нефти (рис. 3.2) могли находиться только в кайнозойском бассейне, т.е. в бассейне того времени, к которому сформировались все известные ныне группы растений и животных.
Химический состав ископаемых осадков в какой-то степени отражает биоценозы, вернее, танатоценозы соответствующих геологических эпох.
В додевонское время основными поставщиками ОВ были синезеленые и прочие водоросли и бактерии, высшие растения отсутствовали, поэтому в отложениях додевонского времени пыльца, кутикула, витринит, растительные воски не наблюдаются. В ОВ этих отложений нет подавляющего превосходства нечетных УВ среди высших нормальных алканов. Сравнивая химический состав ОВ отложений разного возраста (имеется в виду ОВ, не измененное или мало измененное в катагенезе), далеко не всегда можно определить «возрастные» отличия или специфику исходного ОВ, поскольку все планктоногенное ОВ в отложениях различного возраста сформировано за счет фитопланктона и бакте-
|
рий; все остальные поставщики ОВ — незначительная примесь, которая может и не сказаться на химическом составе захороненного ОВ.
Заметные отличия в химическом составе ОВ фиксируются только для двух крайних типов: 1) планктоногенного, образованного за счет жизнедеятельности фитопланктона и бактерий, чисто сапропелевого; 2) органического вещества наземного происхождения, образованного за счет высших растений, гумусового или арконового ОВ. Помимо отличий в углеводородной части меняется валовый химический состав, что приводит, в частности, к изменению величины Н/Сат; для сапропелевого (I тип кероге-на) ОВ она составляет 1,7-1,9, в то время как в более ароматическом обогащенном кислородом гумусовом ОВ ( III тип кероге-на) величина этого отношения не превышает 1,1.
Известно, что климат и температура воды влияют на жизнедеятельность организмов и состав образованных исходным ОВ продуктов. Так, снижение среднегодовых температур, происходящее в связи с похолоданием от кембрия до современного периода, повлияло на структуру масел исходного ОВ в направлении повышения содержания ненасыщенных жирных кислот, присущих данному виду. Поскольку экспериментально установлено, что насыщенные жирные кислоты — источник алканов, а ненасыщенные — нафтеновых УВ, то соотношение алканы/цик-ланы, производных насыщенных и ненасыщеных кислот, синтезируемых растениями, контролируется температурой среды обитания.
Характер распределения некоторых биомаркеров (стеранов С28 и С29) в УВ фракции морского ОВ и нефтей различного возраста в геологической истории позволил наметить их генетическую связь с эволюцией основных продуцентов ОВ (рис. 3.3). Эта эмпирически установленная закономерность — увеличение отношения стеранов С28/С29 от кембрия к кайнозою — не объясняет характер распределения стеранов в докембрийском ОВ и нефтях. На эмпирической кривой докембрийскому ОВ соответствуют низкие значения этого отношения, какие наблюдались в известных ранее нефтях докембрия. В результате исследований последних лет ОВ рифейских отложений и нефтей Восточно-Европейской платформы было идентифицированы ОВ и нафтиды с равномерным распределением стеранов, т.е. отношения стеранов С28/С29 ~ 1, что соответствует ОВ динофлагеллат, появившихся много позже.
До сих пор состав исходных биопродуцентов докембрия изучен крайне слабо. Не исключено, что в докембрии существовали предшественники динофлагеллат, биохимический цикл которых был сходен, что и предопределило соответствующее распределение биомаркеров. Таким образом, данные об углеводородном со-
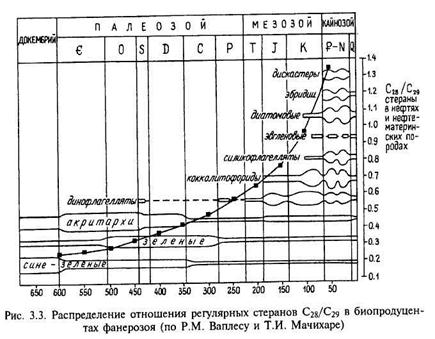
ставе, характере распределения биомаркеров позволяют решать и (или) поставить обратную задачу: каковы же были биопродуценты прошлых геологических эпох, особенно это касается ОВ наиболее древних и менее изученных до кембрийских отложений.
Date: 2015-04-23; view: 1783; Нарушение авторских прав; Помощь в написании работы --> СЮДА... |